
Download
PAPER
Phytochemical component, and antioxidant and vasculo-protective activities of Taiwan cocoa polyphenols by different processing methods
Heuy-Ling Chu1, Hong-Xuan Fu2, En-Kuang Chou2, Ying-Chun Lin1*
1Department of Food Science and Technology, Chia Nan University of Pharmacy and Science, Tainan City, Rende Dist., Taiwan (ROC)
2Department of Pharmacy (with Master’s program), Chia Nan University of Pharmacy and Science, Tainan City, Rende Dist., Taiwan (ROC)
Abstract
Cocoa tree (Theobroma cacao L.) is a recently planted crop in Taiwan, a country located in East Asia. Taiwanese cocoa beans are appreciated globally because of their distinctive flavor and aroma. The effects of the water extracts of unfermented Taiwan cocoa beans (WUFCB) and fermented and roasted Taiwan cocoa beans (WFRCB) on the anti-oxidation, vascular protection, and variation in phytochemical components were investigated. Variations in the catechins components of WUFCB and WFRCB were examined by high performance liquid chromatography. The values of catechin compounds in WUFCB (epicatechin [EC]: 52.32 ± 0.56 mg/g, and catechin (C): 15.14 ± 0.26 mg/g) were approximately two times higher than those found in WFRCB (EC: 26.22 ± 0.48 mg/g, and C: 4.56 ± 0.10 mg/g), indicating that the fermentation and roasting steps caused decline in catechins compounds of WFRCB. In the range of 50–300 µg mL-1, both WUFCB and WFRCB depict noncytotoxicity in endothelial cells; they protect cells from H2O2-induced cytotoxicity as established by MTT assay. Meanwhile, nitric oxide (NO) levels in endothelial cells were elevated by WUFCB. In addition, WUFCB displayed radical scavenging in the acellular model and inhibited increase in reactive oxygen species (ROS) noted in endothelial cell induced by H2O2. Overall, the significant vascular protection of WUFCB is associated with increased NO formation. The decreased ROS generation against oxidative damage was attributed to abundant catechin compounds. This study establishes Taiwan cocoa polyphenol as an effective and complementary tool for preventing endothelial dysfunction and cardiovascular disease.
Key words: endothelial cells, fermentation and roasting, oxidative damage, Taiwan cocoa polyphenol, vascular protection
*Corresponding Author: Ying-Chun Lin, Department of Food Science and Technology, Chia Nan University of Pharmacy and Science, No. 60, Sec. 1, Erren Rd., Tainan City, Rende Dist. 71710, Taiwan (ROC). Email: [email protected]
Received: 28 September 2021; Accepted: 8 February 2022; Published: 25 February 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
The impression of humans toward chocolate products has gradually shifted from desserts to healthcare products, emphasizing the amount of bioactive substances found in cocoa beans. Polyphenols in cocoa beans not only bring out unique astringency and bitterness of chocolate flavor but also possess numerous benefits to human health, for instance, eyesight protection (Puell and de Pascual-Teres, 2021); prevention of Parkinson’s and Alzheimer’s diseases, improved recognition ability, prevention of obesity, increasing the amounts of adiponectin and glucose transporter, decreasing the production of lipid, and insulin resistance (Magrone et al., 2017). Moreover, Buijsse et al. (2010) discovered that polyphenols relax the smooth muscles, having the potential to reduce blood pressure. Heiss et al. (2010) demonstrated that polyphenols prevent the aggregation of platelets and moderate inflammation (Monagas et al., 2009). All these mentioned benefits are related to a critical neurotransmitter in the cardiovascular system, that is, nitric oxide (NO). Nitric oxide serves as a second messenger for signaling the cardiovascular smooth muscles. Small molecular weight and lipophilicity of NO allow it to rapidly pass through cell membranes and reach the smooth muscle cells. Sufficient NO bioavailability is associated with normal vasodilation and normal blood pressure. The factors influencing NO levels are as follows: (1) Most NO is synthesized from L-arginine in a reaction catalyzed by the enzyme of endothelial NO synthase (eNOS). The NO level depends on eNOS expression and activity. (2) NO reacts with superoxide anion (O2-) to form peroxynitrite (ONOO-), which can oxidize cell components, eliminate NO levels and suppress NO bioavailability. (3) The major sources of reactive oxygen species (ROS) (including H2O2, OH. and O2-.) are related to mitochondrial electron transport chain and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) pathway. The elimination of ROS production is associated with decreasing cellular oxidative stress, generation of ONOO- and increased NO levels (Fraga et al., 2011 ; Oteiza et al., 2021). As indicated, cocoa polyphenol is involved in these phenomena.
According to the statistics provided by the International Cocoa Organization (ICCO), global cocoa bean production in 2018–2019 season exceeded 4.78 million tons, and the production of 2019–2020 season exceeded 4.69 million tons (International Cocoa Organization, 2020; Olga et al., 2020). In Taiwan, the planting area of cocoa has increased in recent years, particularly in Neipu and Wanluan counties of Pingtung, Southern Taiwan, where cocoa plantation is done in about 350 hectares. Nowadays, the distribution of cocoa tree planting area has even reached to the middle of Taiwan, indicating cocoa trees have become a recently developed crop in Taiwan. This has led the government and academic institutions to conduct cocoa-relevant research, for instance, the effect of choice of fermentation of starter cocoa cultures, roasting temperature of cocoa beans leading changes in flavanols and procyanidins and sensory properties of cocoa end products. All these investigations aim to enrich the competitiveness and value of Taiwan’s cocoa products. However, few studies are conducted involving the cytoprotection of Taiwan cocoa beans and content levels of catechins and methylxanthine compounds in fermented and unfermented cocoa beans, both of which are expected to have higher phytochemical contents.
The present study was designed to prepare two types of cocoa beans under different processing methods (i.e., fermented and unfermented) to explore the effect of their extracts concerning phytochemical level, antioxidant activity, diminishing oxidative stress and vascular protection of endothelial cells. The study also evaluated function of Taiwan cocoa beans to effectively improve endothelial function.
Material and Methods
Materials
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), epicatechin, catechin, theobromine, caffeine, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA). Mouse vascular endothelial cells, SVEC4-10 (BCRC No. 60220) were purchased from the Bio-resource Collection and Research Center (BCRC, Food Industry Research and Development Institute, Hsinchu, Taiwan). All chemicals of analytical grade were used in the research.
Preparation of cocoa beans
Cocoa pods were purchased from a Pingtung farm in Taiwan. The husk and damaged parts of the cocoa pods were removed. Then 60-kg cocoa beans were fermented for 7 days and sun-dried until the moisture level was 6%. Following fermentation and sun-drying, the cocoa beans were randomly sampled for roasting. The beans were roasted for 25 min at a temperature of 130°C. The final roasted cocoa beans were known as fermented and roasted cocoa beans.
On the other hand, another 20 kg of harvested cocoa beans were in boiling water for 15 min at 90°C and at once cooled in ice water. Then, the boiled cocoa beans were washed and sun-dried until the moisture level dropped to 6%. These bean samples were known as unfermented cocoa beans.
Extraction
A total of 100 g of cocoa beans were peeled manually for each processing method and the nibs were ground into powder in a grinder with liquid nitrogen. The cocoa powder was defatted for 20 min and extracted with boiling water in 1:10 (w/v) ratio. After filtering through Advantec No, 2. filter paper, the residue was reextracted using the same steps. The supernatant was collected and concentrated through a rotary evaporator, freeze-dried, and stored under -20°C. Two different cocoa extracts were obtained, viz. water extracts of fermented and roasted cocoa beans (WFRCB) and water extracts of unfermented cocoa beans (WUFCB).
High-performance liquid chromatography (HPLC) analysis marker components in cocoa extract
The analysis was performed with HPLC according to the method enumerated by Zhang et al. (2016). HPLC chromaster (Hitachi Ltd., Tokyo, Japan) comprised 7725i injector, 5110 pump and 5430 diode array detector (DAD). Sample (1 mg mL-1) was filtered through a hydrophilic PVDF 0.22-µm membrane and injected with a 20-µL sample into an RP-18 column (5µm particle, 4.6 × 250 mm, Mightysil [Kanto Chemical, Tokyo, Japan]). The elution buffer consisted of acetonitrile (Solvent A) and 1% (v/v) phosphoric acid in water (Solvent B) at a flow rate at 0.8 mL/min. The solvent gradient was as follows: 0–8 min, 5–15% B, 8–30 min, 15–25% B, 30–40 min, 25–30% B followed by equilibration of the column prior to new injection. All compounds were detected at 280 nm. Based on the profile of the concentration (x, µg mL-1) versus peak area (y), the linear regression equation and the correlation coefficients (r2) were as follows: theobromine: y = 7,810.1x -14,896 (r2 = 0.997); epigallocatechin (EGC): y = 2,700x -21.0 (r2 = 0.999); caffeine: y = 31,668x +9,043.1 (r2 = 0.999); catechin (C): y = 8,278.9x –1,790.9 (r2 = 0.999); epicatechin (EC): y = 9,274.6x +13,850 (r2 = 0.994); epigallocatechin gallate (EGCG): y = 9,929.2 x -474.1 (r2 = 0.999); and epicatechin gallate (ECG): y = 10,790x -445.4 (r2 = 0.999). The HPLC analysis for each sample was performed in triplicate.
Total polyphenolic content (TPC)
Total polyphenol content was analyzed as gallic acid equivalent (GAE). Different concentrations of cocoa extracts were added in a 10-mL volumetric flask, to which 2-mL sodium carbonate (20% w/v) was added. After 5 min, 0.1-mL Folin–Ciocalteu reagent (50% v/v) was added and the volume was made up to 10 mL with H2O. After 1-h incubation at 30°C, the absorbance was measured at 750 nm and compared to a gallic acid calibration curve.
Proanthocyanidins content (PAC)
The working solution consisted of 0.25-mL cocoa extract, 1.5-mL n-BuOH/HCl (95:5, v/v) and 50-µL 2% solution of NH4Fe(SO4)2 12H2O in 2-M HCl. The reaction mixture was capped, shaken and incubated for 1 h at 95°C. The solution was cooled at room temperature and the absorbance was measured at 550 nm using an enzyme-linked-immunosorbent assay (ELISA) reader (Molecular DevicesVMax, MA, USA). The concentration of proanthocyanidin was measured from a standard curve of cyanidin chloride with the same protocol, and was indicated as mg cyanidin chloride equivalent (CyE/g) of cocoa extract.
DPPH radical scavenging activity
The effect of cocoa extracts on reducing the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity was determined. The samples were added to a methanolic solution (1 mL) of DPPH radicals (final concentration of DPPH was 0.2 mM). The mixture was shaken vigorously for 30 min at room temperature and then placed in dark. Absorbance of the resulting solution was measured at 517 nm. Trolox equivalent (TE) was used as a reference standard.
Trolox equivalent antioxidant capacity (TEAC) assay
The 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS+) cation-free radical inhibitory activity was measured as described previously (Mellinas, et al., 2020). The ABTS+ was generated by reacting 1-mM ABTS+ with 0.5-mM hydrogen peroxide and 10 units/mL horseradish peroxidase in dark at 30°C for 2 h. After addition of 1-mL ABTS+ to cocoa extract, absorbance was measured after 10 min at 734 nm. A lower level of absorbance indicated a stronger inhibitory activity of samples. Trolox equivalent was again used to obtain calibration curve.
SVEC4-10 cell line culture
SVEC cells were obtained from BCRC, Taiwan. The cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% heat-inactivated fetal bovine serum and 1.5 g L-1 L-glutamine at 37°C in a humidified 5% CO2/95% air-controlled incubator.
Cell viability assay
Cell viability was performed by MTT assay. MTT is a tetrazolium salt and was converted into insoluble formazan by mitochondrial succinate dehydrogenase of living cells. Briefly, cells were seeded in 96-well plates cultured for 24 h. Next, the cells were treated with sample for 1 h, H2O2 was added to the medium and incubated for 24 h. Then 5 mg mL-1 MTT stock solution was added in each well and incubated for 2 h. Subsequently, the medium was removed and the plates were incubated for 30 min to solubilize colored formazan dye by the addition of DMSO. Then the absorbance of each well was measured with a Thermo Model 355 microplate reader at 550 nm.
Intercellular reactive oxygen species
In order to determine the generation of ROS in cells, 2’,7’-dichlorofluorescein-diacetate (DCFH-DA) was used. It penetrates cell membranes and is hydrolyzed by intracellular esterase to form dichlorodihydro-fluorescein (DCFH). DCFH reacts with ROS generated by intracellular stress to produce highly fluorescent DCF, which emits fluorescence when excited at 485 nm. The SVEC4-10 cells were cotreated with cocoa extract and 0.2-mM H2O2. Subsequently, DCFH-DA (50 µM) was added to the medium and incubated for 240 min. The treatment medium was removed, and the cells were washed twice with phosphate buffer solution (PBS). The ROS produced from intracellular stress was detected using a Bio-Tek FLX800 microplate fluorescence reader (Winoosky, VT, USA) with an excitation wavelength of 485 nm and an emission wavelength of 530 nm.
Intercellular nitric oxide
Nitrite levels in cell-cultured media, reflecting the intracellular NO synthase activity, were determined by the Griess reaction. Briefly, the cells were cultured for 18 h with cocoa extracts. Then the growth medium was added with the same volume of Griess reagent (1% sulfanilamide in 5% phosphoric acid and 0.1% N-(1-naphthyl)ethylenediamine dihydrochloride in water) at room temperature. Then the absorbance of mixture at 550 nm was detected.
Statistical analysis
Each experiment was performed at least for three times and the average was calculated. Data are expressed as mean ± SD, and ANOVA was conducted using the SPSS software (version 12.0, SPSS Inc., Chicago, IL, USA). When a significant F-ratio (mean square between treatments / mean square within treatments) was obtained (p < 0.05), a post hoc analysis was conducted between groups using Duncan’s multiple range tests. A significant difference between treatments was considered for p < 0.05.
Results and Discussion
Principle phytochemicals present in WUFCB and WFRCB using HPLC
Phytochemicals are plant-derived small molecules possessing multifunctional effects and significantly inhibit oxidative stress. It is worth evaluating the variation in the phytochemical components of Taiwan cocoa beans with different processing procedures. The analysis and separation of the phytochemical components present in WUFCB and WFRCB were performed by HPLC-DAD using an RP-18 analytical column. The qualitative analysis of the phytochemicals utilized linear regression equation of commercial standard compound as described in ‘Methods’ section. Seven principle compounds were identified and quantified, including theobromine (49.57 ± 1.06 mg/g WUFCB and 71.22 ± 1.24 mg/g WFRCB), epigallocatechin (2.22 ± 0.31 mg/g WUFCB and 1.06 ± 0.11 mg/g WFRCB), caffeine ( 5.38 ± 0.07 mg/g WUFCB and 8.90 ± 0.16 mg/g WFRCB), catechin (15.14 ± 0.26 mg/g WUFCB and 4.56 ± 0.10 mg/g WFRCB), epicatechin (52.32 ± 0.56 mg/g WUFCB and 26.22 ± 0.48 mg/ g WFRCB), epigallocatechin gallate (EGCG, 10.78 ± 0.12 mg/g WUFCB and 5.00 ± 0.50 mg/g WFRCB) and epicatechin gallate (ECG, 0.40 ± 0.15 mg/g WUFCB and 0.48 ± 0.05 mg/g WFRCB), by comparison between retention time (RT), UV-Vis spectrum, the characteristic maximum wavelength (λmax), and literature data from commercial standard and samples. Variation in methylxanthine and catechin levels between WUFCB and WFRCB was investigated as shown in Table 1. The data indicated that WUFCB was rich in catechin compounds, including catechin, epicatechin, EGCG, and EGC. The values of catechin compounds present in WUFCB were approximately two times higher than those found in WFRCB, except for ECG. Alternatively, there was a lower content of methylxanthines (theobromine and caffeine) in WUFCB involved in the pretreatment; it was boiled in hot water at 90°C for 15 min for inactivating polyphenoloxidase and peroxidase. In the latter situation, a lot of methylxanthines (theobromine and caffeine) were found soluble in hot water (Febrianto and Zhu, 2020) that retained a lower concentration of methylxanthines present in WUFCB. Theobromine, caffeine, catechin, epicatechin and procyanidins are related to the astringency and bitter taste of cocoa products. The bitter taste of cocoa is largely influenced by the content of theobromine and caffeine, and to a lesser degree by polyphenolic compounds. Additionally, theobromine and caffeine have a physiological stimulatory activity on the central nervous system (CNS) with beneficial health effects on cognition, satiety function, and mood (Letricia et al., 2021).
Table 1. HPLC analysis of principle polyphenols present in WUFCB and WFRCB.
| Compound | Retention time (min) | λmax (nm) | Content (mg/g extract) | |
|---|---|---|---|---|
| WUFCB | WFRCB | |||
| Theobromine | 10.1 | 272 | 49.57 ± 1.06 | 71.22 ± 1.24 |
| EGC | 13.6 | 271 | 2.22 ± 0.31 | 1.06 ± 0.11 |
| Caffeine | 14.9 | 272 | 5.38 ± 0.07 | 8.90 ± 0.16 |
| Catechin | 15.3 | 278 | 15.14 ± 0.26 | 4.56 ± 0.10 |
| Epicatechin | 18.1 | 278 | 52.32 ± 0.56 | 26.22 ± 0.48 |
| EGCG | 18.9 | 275 | 10.78 ± 0.12 | 5.00 ± 0.50 |
| ECG | 26.7 | 276 | 0.40 ± 0.15 | 0.48 ± 0.05 |
The data were indicated with mean ± SD.
EGC: epigallocatechin; EGCG: epigallocatechin gallate; ECG: epicatechin gallate; WUFCB: water extracts of unfermented cocoa beans; WFRCB: water extracts of fermented and roasted cocoa beans.
As described in Table 1, the main polyphenol present in both WUFCB and WFRCB was epicatechin. Current research has found that epicatechin can serve as a free radical scavenger, metal chelator, eNOS activator and arginase inhibitor (Fraga et al., 2011), modulating oxidative stress and contributing to the prevention of cardiovascular diseases. Therefore, epicathchin was used as a reference control in the following assay.
Effect of cocoa extracts on antioxidant activity
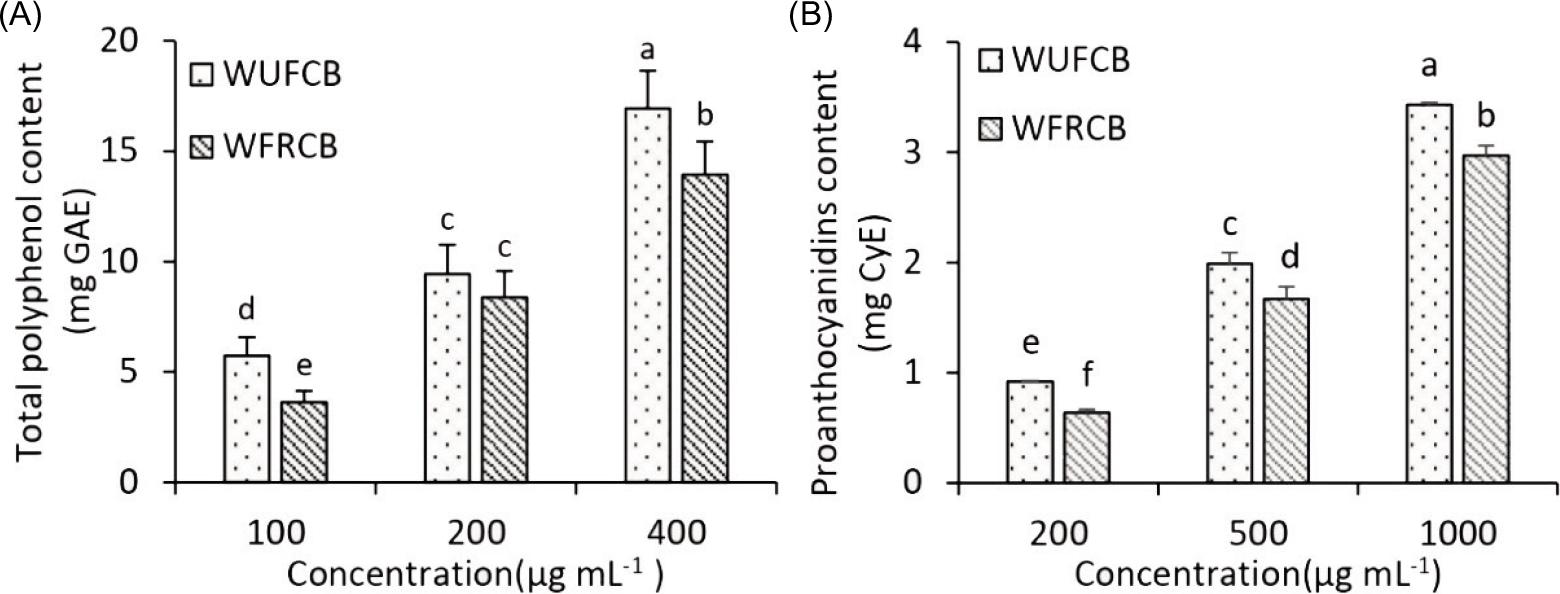
Polyphenol and proanthocyanidins naturally occur in cocoa beans, playing important roles in different biofunctional activities and have positive health effects in humans. The TPC values in WUFCB and WFRCB are shown in Figure 1A, displaying significant difference (p < 0.05). At 400 µg mL-1 of WUFCB and WFRCB, TPC was quantified in 16.93 ± 1.71 mg GAE and 13.84 ± 1.48 mg GAE, respectively. It was suggested that fermentation and roasting of the processing stage significantly decreased TPC, that is, WFRCB is 18.3% lower in TPC after these processing steps.
Figure 1. Determination of (A) total polyphenol content (TPC) and (B) proanthocyanidins content (PAC) in water extracts of unfermented cocoa beans (WUFCB) and water extracts of fermented and roasted cocoa beans (WFRCB). Different superscript letters indicate significant differences (p < 0.05).
Proanthocyanidins have depicted potential beneficial effects on human health, in addition to a pleasant sensory ingredient found in cocoa products (Pedan et al., 2018). The effect of WUFCB and WFRCB on PAC display significant differences, as shown in Figure 1B (p < 0.05). PAC was influenced by fermentation and roasting steps; it decreased from 3.43 ± 0.02 mg CyE/g WUFCB to 2.97 ± 0.09 mg CyE/g WFRCB.
Proanthocyanidins play a potential role in the prevention of cardiovascular disease and this includes the monomer, catechin and epicatechin, and oligomer units. The obtained data from Figures 1A and B confirm the observation that operational process decreased the levels of PAC and TPC. The results were consistent with previous studies, and the following reasons explain the higher levels of PAC and TPC in WUFCB than in WFRCB: (1) WUFCB (unfermented cocoa beans) were raw cocoa beans and had not undergone the process of fermentation and roasting. Polyphenolic compounds diffused from cellular compartments and oxidized to produce insoluble high molecular weight tannins during the fermentation period. These oxidation reactions were catalyzed by polyphenol oxidase (PPO), which could be the remaining 5–10% of enzymatic activity between the first and the second day of fermentation (Febrianto and Zhu, 2020); and (2) the pretreated WUFCB were heated at 90°C for 15 min, which almost completely inhibited PPO activity. Lower PPO activity retained more TPC and PAC. Further, lower PAC and TPC contents in WFRCB could be because these compounds were susceptible to oxidative degradation during fermentation and roasting (Quiroz-Reyes and Fogliano, 2018).
DPPH and ABTS methods, associated with free radical scavenging activities, were selected to assay antioxidant activity. The effect of WUFCB and WFRCB on DPPH and ABTS radical scavenging activities are shown in Table 2. Both WUFCB and WFRCB exhibit a concentration-dependent effect for these two methods in terms of free radical scavenging assay and display a significant difference (p < 0.05). The values of antiradical activity by DPPH were similar to that of ABTS. Although similar results were obtained in both DPPH and ABTS assays, properties of each of the extracted compounds differed. In DPPH assay, it easily interacted with hydrophobic compounds; however, it interacted with both hydrophilic and hydrophobic molecules in ABTS assay (Carmen L.D.T.S. et al., 2021). The DPPH radical scavenging activities for WUFCB and WFRCB at concentrations of 150 µg mL-1 were equal to 9.70 ± 0.82 mg TE and 8.86 ± 0.62 mg TE, respectively. The ABTS radical scavenging activities were 11.81 ± 0.26 mg TE and 9.26 ± 0.39 mg TE, respectively. WUFCB exhibited more antiradical activity than WFRCB. The obtained results demonstrated that more of processing steps decreased both free radical scavenging activities.
Table 2. The effect of WUFCB (water extracts of unfermented cocoa beans), and WFRCB (water extracts of fermented and roasted cocoa beans) on DPPH and ABTS scavenging activity.
| Sample | Concentration (μg mL–1) | DPPH scavenging activity (mg TE) | ABTS scavenging activity (mg TE) |
|---|---|---|---|
| Epicatechin | 50 | 4.37 ± 0.39e | 3.59 ± 0.57e |
| 100 | 5.43 ± 0.49de | 6.82 ± 0.34d | |
| 150 | 6.87 ± 0.90bcd | 10.60 ± 0.31ab | |
| WUFCB | 50 | 7.50 ± 0.85bc | 4.48 ± 0.71e |
| 100 | 8.74 ± 0.94ab | 8.25 ± 0.43c | |
| 150 | 9.70 ± 0.82a | 11.81 ± 0.26a | |
| WFRCB | 50 | 6.68 ± 0.52cd | 3.48 ± 0.65e |
| 100 | 8.09 ± 0.66abc | 6.52 ± 0.53d | |
| 150 | 8.86 ± 0.62ab | 9.26 ± 0.39bc |
The data were indicated with mean ± SD.
The different letters indicated significant differences (p < 0.05).
The data obtained in Figure 1 and Table 2 show that less processing steps implies higher retention of TPC and PAC, and DPPH and ABTS radical scavenging activities. The results also established that the antioxidant capacity is directly related to TPC and PAC, which, of course, are affected by the process of fermentation, drying and roasting. The main polyphenolic compounds present in cocoa beans are epicatechin and PAC, which are composed of aromatic rings and functional groups such as hydroxyl group. These structures allow TPC and PAC to chelate metal ions, resulting in the formation of inactive complexes, or the activity to stabilize and reduce hydroxyl, peroxyl or superoxide radicals (Noelia et al., 2021). Free radicals play a crucial role in causing oxidative damage and pathological events, including inflammation, aging and cardiovascular diseases. These observations demonstrated that antiradical activities were potentially affected by TPC and PAC found in cocoa and the low-processing cocoa extract (i.e., WUFCB), rich in TPC and PAC and higher antiradical activity, was a promising candidate for becoming a potential health-promoting food.
In order to examine whether the antiradical activity is related to TPC and PAC, a correlation coefficient test (r2) was performed. Previous studies have reported a strong correlation between polyphenolic compounds with the antiradical activity of extracts (Carmen L.D.T.S. et al., 2021). Based on the concentration of TPC (x, mg GAE) or PAC (x, mg CyE) versus DPPH (y, mg TE) and ABTS scavenging activities (y, mg TE), the linear regression equation and the correlation coefficient (r2) are as follows: TPC-DPPH /WUFCB: y = 1.1383x + 5.007 (r2 = 0.992); TPC-ABTS/WUFCB: y = 2.5858x - 5.6934 (r2 = 0.999); PAC-DPPH/WUFCB: y = 12.108x + 3.889 (r2 = 0.990); PAC-ABTS/WUFCB: y = 31.462x - 10.461 (r2 = 0.998); TPC-DPPH/WFRCB: y = 1.6322x + 4.1053 (r2 = 0.987); TPC-ABTS/WFRCB: y = 1.9505x - 0.0044 (r2 = 0.965); PAC-DPPH/WFRCB: y = 16.93x + 3.4375 (r2 = 0.996); PAC-ABTS/WFRCB: y = 15.483x + 0.6961 (r2 = 0.972).
The high correlation values observed for antiradical activity are related to both TPC and PAC. The r2 varied from 0.965 to 0.999.
Effect of cocoa extracts on cell viability in endothelial cells
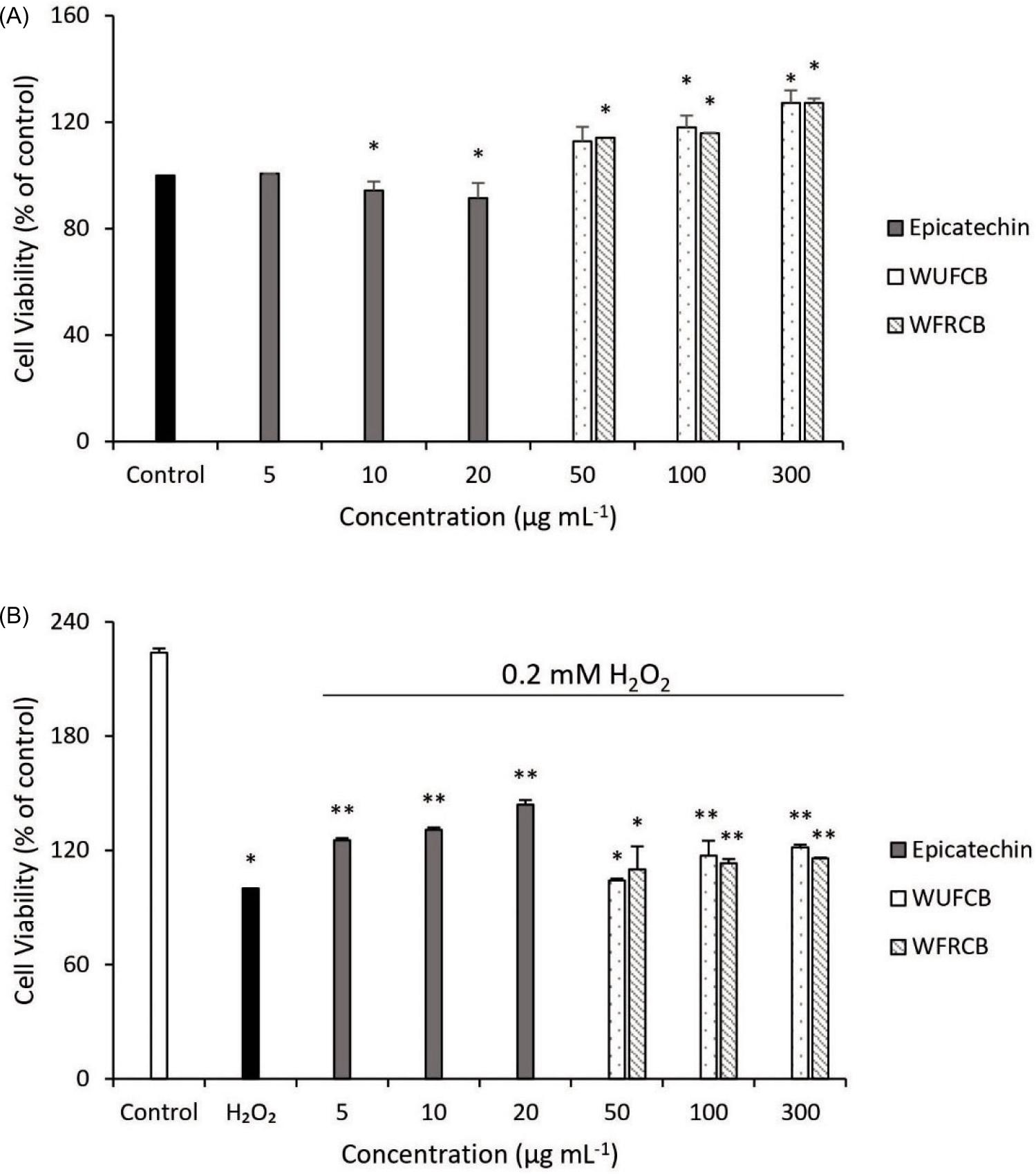
Endothelial cells are susceptible to oxidative damage, resulting in disturbed cardiovascular homeostasis. For this reason, the endothelial cell line SVEC4-10 was used in this study. MTT assay is widely used to measure cell growth. The cytotoxicity of cocoa extract and epicatechin was determined by an MTT assay. Figure 2A shows the response of endothelial cells incubated with samples for 24 h. The cell viabilities of two cocoa extracts of 50–300 µg mL-1 and epicatechin at 5–20 µg mL-1 were from 91.46% to 127.31% compared to untreated control cells. In other words, WUFCB, WFRCB and epicatechin demonstrated non-cytotoxicity in endothelial cells at all studied concentrations.
Figure 2. Cell viability relative to (A) untreated control and (B) damage of endothelial cells by 0.2-mM H2O2 following different concentrations of water extracts of unfermented cocoa beans (WUFCB), water extracts of fermented and roasted cocoa beans (WFRCB) and epicatechin, estimated with MTT assay. The data were indicated as mean ± SD. Different markers indicate significant differences (p < 0.05).
Effect of cocoa extracts on cytoprotection by H2O2-induced oxidative stress in endothelial cells
Free radicals, including O2-.,OH., and H2O2, are toxic to vascular endothelial cells. H2O2 has no charge and can diffuse through the cell membrane. Therefore, in this research H2O2 was selected as an oxidant reagent to induce oxidative stress in endothelial cells. In order to evaluate whether Taiwan cocoa polyphenol was involved in protecting endothelial cells from H2O2-induced oxidative stress, direct cytotoxicity effect of H2O2 on endothelial cells was examined by MTT assay. This included 50, 100 and 300 µg mL-1 of cocoa extracts and 5, 10 and 20 µg mL-1 of epicatechin. Endothelial cells were damaged by H2O2 for 1 h, and the samples were added and incubated for 24 h. The viability of untreated or treated cells by 0.2-mM H2O2 in the absence of samples was 223.85% and 100%, indicating that H2O2 demonstrated significant cytotoxicity on endothelial cells (Figure 2B). Treatment of 300 µg mL-1 of WUFCB and WFRCB and 20 µg mL-1 of epicatechin with 0.2-mM H2O2 resulted in increased survival rates of endothelial cells, viz. 121.52%, 115.75% and 143.97%, respectively, compared with untreated cells (100%). The results in Figure 2B indicated the cytoprotective effects of WUFCB, WFRCB, and epicatechin from H2O2-induced cytotoxicity and depicted a dose-dependent increase in cell growth. In this study, epicatechin, a major component of Taiwan cocoa beans, acted as an effective free radical scavenger in acellular and cellular models, and modulated oxidative stress.
Effect of cocoa extracts on ROS generation in endothelial cells
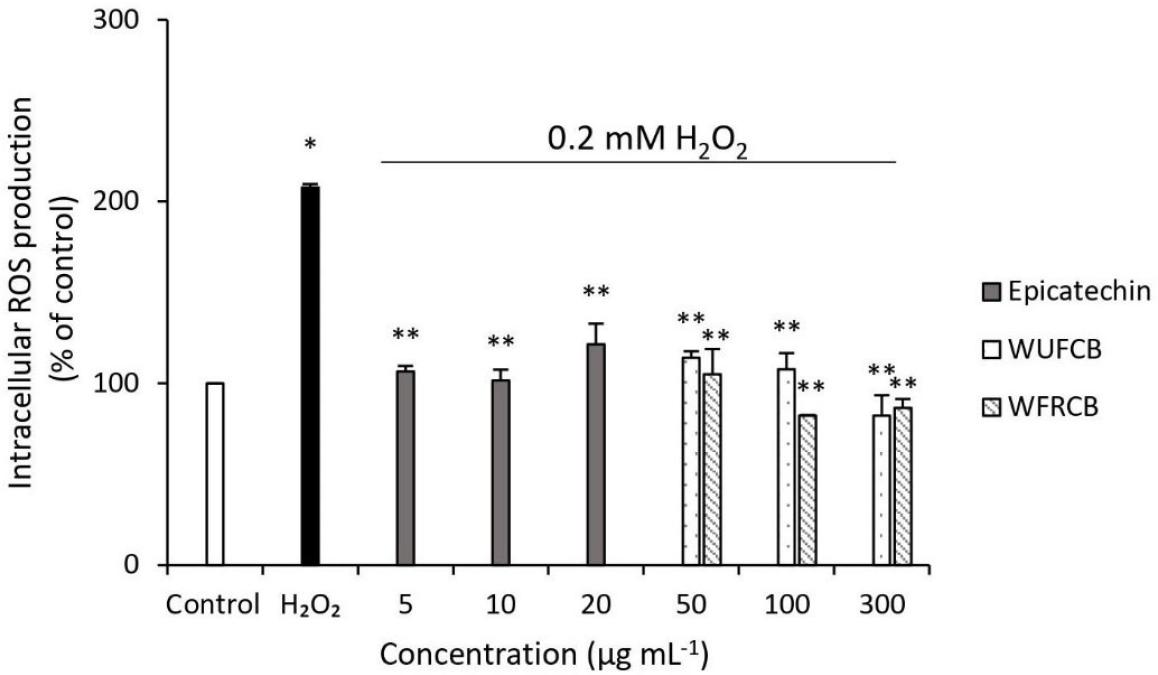
Many studies have revealed that the development of endothelial dysfunction and cardiovascular disease displayed altered redox status compared with normal cells, including increased ROS generation. Additionally, accumulating studies reported that natural phytochemicals, such as EGCG, EGC, ECG, and proanthocyanin, demonstrate a preventive effect, and are involved in ameliorating intracellular accumulation of ROS. To verify whether the cytoprotection of Taiwan cocoa polyphenol was ascribed to the amelioration of oxidative stress, the effects of the WUFCB and WFRCB were examined on the intracellular ROS formation of SVEC4-10 damaged by H2O2. Figure 3 displays the effect of WUFCB, WFRCB and epicatechin on ROS formation in endothelial cells. The intensity of florescence DCF was determined in endothelial cells pretreated with samples for 1 h and then exposed to 0.2-mM H2O2. If the endothelial cells were damaged by H2O2 for 30 min, intracellular ROS increased to 207.8% compared with the corresponding control value (100%). In addition, pretreatment of endothelial cells with 300 µg mL-1 of WUFCB and WFRCB, and 5 µg mL-1 of epicatechin, together with H2O2, decreased intracellular ROS to 82.31%, 86.41% and 106.58%, respectively (Figure 3). There was a statistically significant decrease in ROS formation, indicating that WUFCB, WFRCB and epicatechin significantly ameliorated intracellular accumulation of ROS. Moreover, the identification of catechins compounds by HPLC-DAD, such as epicatechin, against oxidative damage induced by H2O2 may help researchers to better understand protective association of WUFCB with oxidative stress induced by H2O2. This demonstrated that Taiwan cocoa polyphenol could be expected to protect endothelial cells against ROS formation by H2O2-induced oxidative damage.
Figure 3. Effect of water extracts of unfermented cocoa beans (WUFCB), water extracts of fermented and roasted cocoa beans (WFRCB) and epicatechin on intercellular ROS formation in endothelial cells induced by 0.2-mM H2O2. ROS formation is relative to untreated control of endothelial cells and those treated with different concentrations. The data were indicated as mean ± SD. Different markers indicate significant differences (p < 0.05).
Effect of cocoa extracts on NO production in endothelial cell
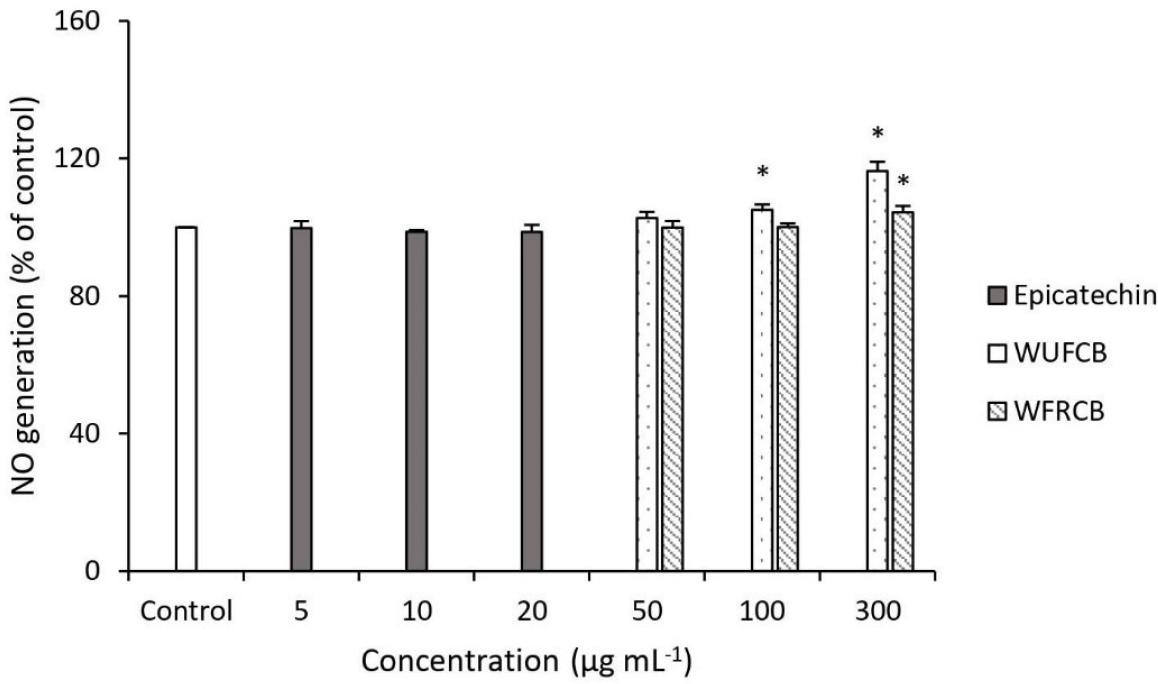
Endothelial nitric oxide synthase produces NO, a freely diffusible gas, acting as a second messenger for signaling vascular smooth muscle relaxation, causing vasodilation and lowering of blood pressure. However, a fall in NO steady-state level results in endothelial dysfunction, failure of smooth muscle relaxation, and consequent hypertension (Engler and Engler, 2004; Fraga, et al., 2011). In this regard, we determined whether WUFCB, WFRCB and epicathchin modulated NO generation in endothelial cells. NO is a short-lived free radical that rapidly reacts with molecular oxygen to yield nitrogen dioxide, dinitrogen trioxide and nitrite. Nitrite is the only stable product. Nitrite levels in the culture medium were determined as an index of NO synthesis by using the Griess reagent (Chu et al., 2017). Incubation with samples for 18 h demonstrated that epicatechin had no significant effect on NO generation (Figure 4). Alternatively, treatment of endothelial cells with 300-µg mL-1 WUFCB resulted in 116.38% NO generation compared with the untreated cells (100%). In other words, WUFCB has the potency to increase NO generation in endothelial cells. This result is in agreement with previous reports (Engler and Engler, 2004) that demonstrated that polyphenol-rich cocoa could activate eNOS for synthesis NO, inhibit arginase and NADPH oxidase, leading to lower levels of O2-.; it could also raise the level of NO (Giovanni et al., 2014). Moreover, tetramers and higher polymers of procyanidin were related to the activation of eNOS. According to the analysis of WUFCB using HPLC-DAD (data not shown), the content of tetramer and higher degree of polymerization (DP) procyanidins were abundant in WUFCB.
Figure 4. Effect of water extracts of unfermented cocoa beans (WUFCB), water extracts of fermented and roasted cocoa beans (WFRCB) and epicatechin on intercellular NO production in endothelial cell. NO production is relative to untreated control of endothelial cells and those treated with different concentrations. The data were indicated as mean ± SD. Different markers indicate significant differences (p < 0.05).
Conclusion
This study demonstrated the basis for the exploitation of polyphenolic compounds extracted from Taiwan cocoa beans. The results provided profiles of polyphenol compounds of WFRCB compared to WUFCB. WUFCB is rich in catechins compounds; additionally, WUFCB exerts a marked vascular protective activity in endothelial cells induced by H2O2 that could be attributed to the higher levels of polyphenols, such as epicatechin, catechin, EGC, EGCG and other compounds, present in WUFCB. These exert their protective action against oxidative stress through scavenging ROS, and modulate NO generation. Therefore, WUFCB has a great potential for preventing endothelial dysfunction and related complications.
Declaration of Interest
The authors declared no conflicts of interest.
Acknowledgments
This work was funded by the Ministry of Science and Technology of the Republic of China [MOST 108-2221-E-041-001].
Contribution of Authors
Ying-Chun Lin designed the study and Heuy-Ling Chu supervised data collection. Hong-Xuan Fu and En-Kuang Chou analyzed the data. Heuy-Ling Chu and Ying-Chun Lin interpreted the data and reviewed the manuscript for publication. All authors approved the final manuscript.
REFERENCES
Buijsse B., Weikert C., Drogan D., and Bergmann M. 2010. Chocolate consumption in relation to blood pressure and risk of cardiovascular disease in German adults. Eur Heart J. 31(13):1616–1623. 10.1093/eurheartj/ehq068
Carmen L.D.T.S., Francisco R.F., Francisco J.C.M., Josue’ J., Sau’l R.C., Francisco J.W.C., Jesu’s B.F., Daniela D.C., Carlos G.B.U., Jose’ A.T.H. 2021. Recovery of phytochemical from three safflower (Carthamus tinctorius L.) by-products: antioxidant properties, protective effect of human erythrocytes and profile by UPLC-DAD-MS. J Food Process Preserv. 45(9):e15765. 10.1111/jfpp.15765
Chu C.C., Wu W.S., Shieh J.P., Chu H.L., Lee C.P., and Duh P.D. 2017. The anti-inflammatory and vasodilating effects of three selected dietary organic sulfur compounds from allium species. J Funct Biomat. 8(1):5. 10.3390/jfb8010005
Engler M.B. and Engler M.M. 2004. The vasculoprotective effects of flavonoid-rich cocoa and chocolate. Nutr Res. 24(9):695–706. 10.1016/j.nutres.2004.05.001
Febrianto N.A. and Zhu F. 2020. Changes in the composition of methylxanthines, polyphenols, and volatiles and sensory profiles of cocoa beans from the Sul 1 genotype affected by fermentation. J Agric Food Chem. 68(32):8658–8675. 10.1021/acs.jafc.0c02909
Fraga C.G., Litterio M.C., Prince P.D., Calabró V., Piotrkowski B., and Galleano M. 2011. Cocoa flavanols: effects on vascular nitric oxide and blood pressure. J Clin Biochem Nutr. 48(1):63–67. 10.3164/jcbn.11–010FR
Giovanni S., Sergio D., Laura D.R., Antonino De L., Hector H.O., Giuseppe M., Arrigo F.C., and Salvador G. 2014. Cocoa bioactive compounds: significance and potential for the maintenance of skin health. Nutrients. 6(8):3202–3213. 10.4145303/pdf/nutrients-06-03202
Guillermo S., Mosca S., Cienfuegos-Jovellanos E., Pasamar M.A., Muguerza B., Ramón D. and Ríos J.L. 2010. Antioxidant properties of polyphenol-rich cocoa products industrially processed. Food Res Int. 43:20101614. 10.1016/j.foodres.2010.04.032
Heiss C., Keen C.L., and Keml M. 2010. Flavanols and cardiovascular disease prevention. Eur Heart J. 31(21):2583–2592. 10.1093/eurheartj/ehq332
International Cocoa Organization (ICCO). 2020, Dec 02. The Quarterly Bulletin of Cocoa Statistics, Vol. XLVI, No. 4, Cocoa year 2019/2020. The International Cocoa Organization, Abidjan, Ivory Coast. Available at: https://www.icco.org/november-2020-quarterly-bulletin-of-cocoa-statistics/
Letricia B.P., Simona B., Ilario F., Olga R.P., and Giuseppe Z. 2021. Characterization and classification of cocoa bean shells from different regions of Venezuela using HPLC-PDA-MS/MS and spectrophotometric techniques coupled to chemometric analysis. Foods. 10:1791. 10.3390/foods10081791
Magrone T., Russo M.A., and Jirillo E. 2017. Cocoa and dark chocolate polyphenols: from biology to clinical applications. Front Immunol. 9(8):677. 10.3389/fimmu.2017.00677
Mellinas, A.C., Jimenez, A. and Garrigós M.C. 2020. Optimization of microwave-assisted extraction of cocoa bean shell waste and evaluation of its antioxidant, physicochemical and functional properties. LWT-Food Science and Technology. 127:1–13. 10.1016/j.lwt.2020.109361
Monagas M., Khan N., Andres-Lacueva C., Casas R., Urpí-Sardà M., and Llorach R. 2009. Effect of coca powder on the modulation of inflammatory biomarkers in patients at high risk of cardiovascular disease. Am J Clin Nutr. 90:1144–1150. 10.3945/ajcn.2009.27716
Noelia D.M., Mónica C. S., Gonzalo S.M., Dolores R.P., and Pilar M. 2021. Cytotoxicity study of bakery product melanoidins on intestinal and endothelial cell lines. Food Chem. 343:128405. 10.1016/j.foodchem.2020.128405
Olga R.P., Letricia B.P., Giuseppe Z. and Caroline S. 2020. Cocoa bean shell—a by-product with nutritional properties and biofunctional potential. Nutrients. 12(4):1123. 10.3390/Fnu12041123
Oteiza P.I., Fraga C.G., and Galleano M. 2021. Linking biomarkers of oxidative stress and disease with flavonoid consumption: from experimental models to humans. Redox Biol. 42:101914. 10.1016/j.redox.2021.101914
Pedan V., Weber C., Do T., Fischer N., Reich E., and Rohn S. 2018. HPTLC fingerprint profile analysis of cocoa proanthocyanidins depending on origin and genotype. Food Chem. 267:277–287. 10.1016/j.foodchem.2017.08.109
Puell M.C. and de Pascual-Teres S. 2021. The acute effect of cocoa and red-berries on visual acuity and cone-mediated dark adaptation in healthy eyes. J Funct Foods. 81:104435. 10.1016/j.jff.2021.104435
Quiroz-Reyes C.N. and Fogliano V. 2018. Design cocoa processing towards healthy cocoa products: the role of phenolics and melanoidins. J Funct Foods. 45:480–490. 10.1016/j.jff.2018.04.031
Zhang Y.N., Yin J.F., Chen J.X., Wang F., Du Q.Z., Jiang Y.W., and Xu Y.Q. 2016. Improving the sweet after-taste of green tea infusion with tannase. Food Chem. 192:470–476. 10.1016/j.foodchem.2015.07.046