
Download

Original Article
Biogenic amine–producing Enterococcus spp. from anchovy (Engraulis encrasicolus): Isolation and molecular insights
Gamze Ergezen1, Gülsüm Atasoy1, Pınar Şanlıbaba1, Nilüfer Vural2, Rahmi Ertan Anlı1*
1Faculty of Engineering, Department of Food Engineering, Ankara University 06830, Ankara, Türkiye;
2Department of Traditional, Complementary and Integrative Medicine, Biotherapeutic Products Research and Development Program, Ankara Yıldırım Beyazit University, Institute of Public Health 06010, Ankara, Türkiye
Abstract
Biogenic amines (BAs) constitute an important chemical hazard in fish and fishery products, primarily due to their adverse effects on human health. While histamine-producing bacteria have been extensively investigated, the contribution of Enterococcus species to BA formation in fresh marine fish remains poorly characterized. The present study aimed to isolate and identify Enterococcus spp. from fresh anchovy (Engraulis encrasicolus) and to evaluate their capacity to produce major BAs. A total of 56 fresh anchovy samples were analyzed, yielding 24 Enterococcus isolates that were characterized by phenotypic methods and 16S rRNA gene sequencing. The BA production capacities of the isolates were quantified via HPLC following incubation in a standardized liquid screening medium under optimized laboratory conditions. Fourteen isolates (58.33%) exhibited aminogenic activity and were identified as Enterococcus faecalis (42.86%), E. faecium (28.57%), E. lactis (21.43%), and E. durans (7.14%). Tyramine, histamine, tryptamine, cadaverine, and putrescine were detected, with maximum concentrations reaching 307.638, 257.939, 236.928, 165.835, and 56.317 mg/L, respectively. The highest total BA levels, E. faecium BLK18, exhibited the highest levels, reaching 493.547 mg/L. These findings demonstrate that fresh anchovies may serve as a reservoir for aminogenic Enterococcus spp. and highlight their potential role in BA accumulation beyond traditionally implicated bacteria. The results support the inclusion of Enterococcus species in routine monitoring programs for fresh fish products and underline the need for broader, evidence-based strategies to manage BA-related hazards in seafood.
Key words: Enterococcus, fresh anchovy, biogenic amine, food safety, molecular characterization
*Corresponding Author: Rahmi Ertan Anli, Faculty of Engineering, Department of Food Engineering, Ankara University 06830, Ankara, Türkiye
Academic Editor: Prof. Mariella Calasso (SIMTREA)—University of Bari, Italy
Recieved: 2 February 2026; Accepted: 20 April 2026; Published: 14 May 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Biogenic amines (BAs) are biologically active -nitrogen- containing compounds that originate mainly from the microbial decarboxylation of free amino acids or, less frequently, from the amination of aldehydes and ketones. These reactions are catalyzed by amino acid decarboxylase enzymes produced by various microorganisms, -particularly during food processing and storage (Li et al., 2025; Maddaloni et al., 2021; Visciano et al., 2020). Depending on their origin, BA-forming enzymatic activities may be classified as endogenous, arising from the raw material itself, or exogenous, resulting from microbial metabolism during processes such as fermentation, maturation, salting, or marination (In et al., 2024; Ucar and Ozogul, 2024).
Among foodborne microorganisms, both Gram-negative bacteria (e.g., Morganella, Klebsiella, Proteus, Enterobacter) and Gram-positive bacteria, particularly lactic acid bacteria (LAB), have been implicated in BAs production (Buňka et al., 2013; Maddaloni et al., 2021). LAB genera such as Enterococcus, Lactobacillus, Lactococcus, and Pediococcus are of special concern because of their strong decarboxylation potential and their ability to survive under adverse conditions (Buňka et al., 2013; Shanab et al., 2025). The genus Enterococcus exhibits substantial intra-species and inter-species variability regarding the qualitative and quantitative biosynthesis of a broad spectrum of Bas (Zhang et al., 2022). While the capacity to generate these -low-molecular-weight nitrogenous compounds is a widespread trait across the genus, Enterococcus faecalis and Enterococcus faecium are particularly distinguished by their potent tyraminogenic activity (Akpomie et al., 2022). This metabolic dominance is primarily attributed to the prevalent occurrence of the tdc (tyrosine decarboxylase) gene cluster, which often exhibits higher transcriptional activity in these species compared to other members such as E. durans or E. hirae Specifically, E. faecalis tends to accumulate significantly higher concentrations of tyramine in proteinaceous matrices, frequently exceeding the critical toxicological threshold of 100 mg/kg (Connil et al., 2002). Beyond tyramine, the aminergic repertoire of enterococci extends to the production of diamines and polyamines. The synthesis of putrescine via the agmatine deiminase (AGDI) pathway represents a distinguishing taxonomic feature, with E. faecalis being more frequently associated with elevated putrescine levels than its counterparts. Furthermore, while less common, certain strains possess the enzymatic machinery (lysine decarboxylase) required for cadaverine formation, often linked to specific environmental stress responses (Sivamaruthi et al., 2021). The presence of polyamines such as spermidine and spermine, though partly endogenous to the food matrix, can also be modulated by enterococcal metabolic flux, further complicating the chemical profile of the product. In contrast to the relatively consistent tyramine production, the histamine-forming capacity within this genus is considered highly strain-specific, often depending on the horizontal gene transfer of the hdc (histidine decarboxylase) locus (Banicod et al., 2025). This multi-amine variability is of paramount importance for food safety, as histamine and tyramine are regarded as the most hazardous BAs due to their severe toxicological effects. Histamine is the primary causative agent of scombroid fish poisoning, while tyramine is linked to the “cheese reaction” characterized by hypertensive crises and neurological symptoms (Shanab et al., 2025). Although scombroid poisoning is traditionally associated with members of the Scombridae family, increasing evidence indicates that non-scombroid species, including anchovy (Engraulis encrasicolus), may also accumulate hazardous histamine levels under improper storage conditions (Arulkumar et al., 2023). Furthermore, non-toxic amines such as putrescine, and cadaverine may potentiate histamine toxicity by inhibiting detoxification pathways or facilitating intestinal absorption (Dara et al., 2025).
Fish and fishery products are especially susceptible to BA accumulation due to their high content of free amino acids, favorable water activity, and rapid post-mortem biochemical changes (Gao et al. 2023; Muhtar et al. 2025). Inadequate storage conditions, microbial proliferation, and enzymatic activity accelerate the formation of BAs such as histamine, tyramine, tryptamine, putrescine, and cadaverine, which are commonly used as indicators of fish freshness and hygienic quality (Ding and Li, 2024; Visciano et al., 2020). Unlike fresh fish, where BAs are typically absent or present at negligible levels, their accumulation reflects microbial spoilage and inappropriate handling practices (Bayesen and Yüksel, 2023; Fadhlaoui-Zid et al., 2012). Anchovy is one of the most economically and nutritionally important pelagic fish species in the Black Sea region and constitutes a substantial proportion of Türkiye’s total marine fish catch. Due to its high protein and free amino acid content, anchovy represents a suitable substrate for BA-producing microorganisms. While BA accumulation in fermented and processed fish products has been extensively documented (Kočar et al., 2021; Ma et al., 2021; Meng et al., 2022; Mutalipassi et al., 2024; Özyurt et al., 2022; Pons-Sánchez-Cascado et al., 2005; Visciano et al. 2012; Zeng et al., 2013; Zhang et al., 2025), research focusing on the aminogenic microbiota of fresh anchovy remains notably scarce. Most existing studies emphasize BA levels in matured or salted products (Al Bulushi et al., 2009; Pons-Sánchez-Cascado et al., 2005), often overlooking the early microbiological determinants that emerge before technological processing exerts selective pressure on the microbial community. Furthermore, much of the published literature primarily addresses the role of Gram-negative spoilage bacteria as the principal histamine producers (Al Bulushi et al., 2009; Visciano et al., 2020; Ding and Li, 2024). In contrast, the aminogenic potential of Gram-positive LAB, particularly Enterococcus spp. naturally present in marine fish post-harvest, has received considerably less attention (Buňka et al., 2013; Maddaloni et al., 2021).
Many current evaluations also tend to quantify BA concentrations at the product level without establishing a direct mechanistic link to the specific microbial producers involved (Çakmak and Sancak, 2023; Dankar et al., 2025; Javaid et al., 2025; Yilmaz 2024). Such an approach limits the identification of early microbial risk factors during the critical stages of handling and storage (Meng et al., 2022). By characterizing aminogenic Enterococcus strains at the species level in fresh anchovy, the present study provides novel insights into the origin and dynamics of BA formation. This molecular characterization contributes original data essential for developing preventive food safety strategies and improving fish quality monitoring frameworks (EFSA, 2011; Shanab et al., 2025).
This study distinguishes itself from previous literature by transitioning from conventional product-based BA analysis toward a high-resolution, micro-organ-ism-oriented assessment of aminogenic potential. While the presence of Enterococcus spp. in various food matrices is documented, empirical data regarding the occurrence, decarboxylase activity, and molecular characteristics of specific strains isolated from fresh anchovy remain remarkably scarce. The novelty of this research is established through an integrated tri-layered approach that combines phenotypic screening for diverse amino acid decarboxylase activities with precise molecular identification of the isolates and strain-level quantification of BAs via high-performance liquid chromatography (HPLC). By synthesizing these methods, this study establishes a direct, quantifiable link between bacterial identity and aminogenic behavior in a highly perishable seafood matrix. Elucidating these functional properties is not only a contribution to microbial taxonomy but is crucial for refining microbial risk assessments and designing targeted preventive strategies to uphold fish quality and consumer safety. Consequently, the primary objectives were to: (i) isolate autochthonous enterococcal strains from fresh anchovy harvested in Türkiye, (ii) characterize their histidine, lysine, tryptophan, -tyrosine, and ornithine decarboxylase profiles, (iii) perform molecular identification of the BA-producing strains, and (iv) provide a definitive quantification of the BAs produced using HPLC.
Materials and Methods
Collection of samples
A total of 56 fresh anchovy (Engraulis encrasicolus) samples were collected from local landing points in the Middle Black Sea during the 2024–2025 fishing season. The sampling period coincided with the official commercial fishing window, which commenced on September 1, 2024, and concluded on April 15, 2025. All specimens were selected to meet the minimum legal conservation size of 9 cm in total length to ensure representative sampling of the commercial catch. The fresh anchovy samples used in this study had an average weight ranging from 12.50 to 13.85 g. Samples were randomly purchased from retail fish markets located in Ankara, Türkiye. To obtain a representative range of fresh anchovy samples, market-specific information was not recorded.
No heat treatment was applied to the samples to preserve the native microbiota for bacterial isolation. All samples were transported to the laboratory in insulated containers under refrigerated conditions (<4°C). Samples were not frozen at any stage. Upon arrival at the laboratory, anchovies were handled aseptically and processed on the same day without storage.
Isolation and biochemical identification of Enterococcus spp
Prior to microbiological examination, anchovy specimens were aseptically eviscerated by manually removing the heads, scales, and bones. The remaining muscle tissues were finely comminuted to serve as the analytical matrix. To isolate enterococci, a 10 g aliquot of this prepared tissue was transferred into 90 mL of sterile physiological saline (0.85% NaCl; Merck, Germany) and processed using a stomacher (Seward 400, USA) for 10 minutes. The resulting suspension was allowed to equilibrate at room temperature for 20 minutes to facilitate optimal microbial dispersion before further analysis. Serial tenfold dilutions were prepared in sterile saline up to 10–5, and 100 µL aliquots were spread onto Kanamycin Aesculin Azide (KAA) agar (Merck, Germany). Plates were incubated at 37°C for 18–24 h. After incubation, three presumptive Enterococcus colonies exhibiting typical black coloration on KAA agar were randomly selected from each plate for further analysis.
The presumptive Enterococcus isolates were subjected to a systematic phenotypic characterization. All isolates were initially screened for Gram reaction and catalase activity. Following the confirmation of Gram-positive, catalase-negative cocci, further physiological assessments were performed, including the ability to hydrolyze bile esculin and grow in the presence of 6.5% NaCl. Additionally, growth performance was evaluated at 10°C and 45°C to verify the characteristic thermal tolerance of the genus (Goa et al., 2022). These conventional biochemical profiles were used to supplement the molecular identification results obtained through 16S rRNA sequencing. Enterococcus isolates were enriched in Tryptic Soy Broth (TSB; Merck, Germany), whereas reference strains were cultivated in Brain Heart Infusion (BHI) broth (Merck, Germany) and TSB, with all cultures incubated at 37°C for 24 h. For long-term viability, isolates were preserved at −20°C in TSB supplemented with 30% (v/v) glycerol (Merck, Germany). The reference strains (Enterococcus faecalis ATCC 29212, Escherichia coli LMG 3083 (ETEC), and Staphylococcus aureus ATCC 6538) were sourced from the culture collection of the Food Microbiology Laboratory, Department of Food Engineering, Faculty of Engineering, Ankara University. These reference strains were specifically employed to function as positive and negative biological controls across the array of biochemical identification assays.
In vitro assessment of ba production by Enterococcus isolates
The ability of Enterococcus isolates to produce BAs was evaluated in vitro by targeting tyramine, putrescine, cadaverine, histamine, and tryptamine, which are frequently associated with enterococci in food matrices. Amino acid decarboxylase activity of the bacterial isolates was evaluated using a decarboxylase broth formulated according to the method of Bover-Cid and Holzapfel (1999). The composition of the medium is summarized in Table 1. The amino acid standards were obtained from Sigma-Aldrich (St. Louis, MO, USA), whereas all remaining reagents were supplied by Merck (Germany). In all assays, a medium without amino acid supplementation served as a negative control, whereas histamine producer Morganella morganii subsp. -morganii ATCC 25830 (Thermo Scientific, Germany) was employed as the positive control strain. Actively growing cultures were standardized to an optical density of 0.50 at 600 nm (OD600), and 0.1 mL aliquots were inoculated into the respective decarboxylase broths. Inoculated tubes were incubated statically at 30°C for 4–5 days and inspected daily for chromatic variation. The absence of color change (yellow) in control tubes confirmed negative decarboxylation activity, whereas a transition from yellow to purple in amino acid–supple mented media was interpreted as a positive decarboxylation reaction, indicating BA formation, in accordance with the criteria described by Bover-Cid and Holzapfel (1999).
Table 1. Composition of decarboxylase media (modified from Bover-Cid and Holzapfel, 1999).
| Component | g/L |
|---|---|
| Tryptone | 5 |
| Yeast extract | 5 |
| Meat extract | 5 |
| Sodium chloride | 0.25 |
| Pyridoxal-5-phosphate | 0.005 |
| Dextrose (1 g/L) | 1 |
| Bromcresol purple | 0.01 |
| Distilled water | 1 L |
*Precursor amino acids were separately incorporated into the basal medium at a final concentration of 0.5% (w/v).
**The pH of the media was adjusted to 5.3 ± 0.02 using sterile 1 N HCl or 1 N NaOH before autoclaving.
***Media were autoclaved at 121°C for 10 min, and separate broth tubes were prepared for each amino acid.
Genotypic identification of Enterococcus spp
BA-producing Enterococcus isolates were genetically identified through amplification and sequencing of the 16S rRNA gene. Genomic DNA was extracted from overnight cultures grown in TSB using a commercial genomic DNA isolation kit (Brand: GeneAll Biotechnology, Catalog No. 106–101, Korea), -following the manufacturer’s protocol. DNA yield and purity were assessed spectrophotometrically using a NanoDrop ND-2000 instrument (Thermo Scientific, USA). Purified DNA samples were subsequently stored at −20°C until further analysis. To ensure the validity of the polymerase Chain Reaction (PCR) assays, each run included a negative control (DNA-free) and appropriate positive control strains. PCR amplifications were performed with little modification as previously described by Beasley and Saris (2004). Amplification of the 16S rRNA gene was performed using the universal primer pair 27F (5´ AGA GTT TGA TCC TGG CTC AG 3´) and 907R (5´ CCG TCA ATT CCT TTG AGT TT 3´), and amplified 900-bp fragments were harvested. PCR were conducted in a final volume of 50 μL, consisting of 3 μL of genomic DNA template, 34.75 μL RNase/ DNase-free water, 5 μL 1×PCR reaction buffer, 4 μL of 25 mM MgCl2, 1 μL of 2 mM dNTPs, 1 μL forward and reverse primers, and 0.25 μL Taq DNA polymerase. The reactions were carried out in a thermal cycler (Techne TC-512, Staffordshire, UK). Thermal cycling conditions included an initial denaturation step at 95°C for 4 min, followed by 30 amplification cycles of denaturation at 95°C for 30 s, primer annealing at 55°C for 30 s, and elongation at 72°C for 6 min. A final extension step was performed at 72°C for 8 min to ensure complete amplification. PCR amplicons were purified using the GeneJET PCR Purification Kit (Thermo Scientific, USA) and resolved by electrophoresis on 1% (w/v) agarose gels prepared in 1× TAE buffer (Thermo Scientific, USA). Following staining with 0.5 μg/mL ethidium bromide (Sigma-Aldrich, USA) for nucleic acid staining DNA bands, electrophoresis was performed at 90–100 V for approximately 45–60 min, and the DNA bands were visualized under UV illumination. Fragment sizes were estimated using an O’GeneRuler™ 10,000 bp DNA ladder (Thermo Scientific, USA). Purified PCR products were sequenced using the Sanger sequencing method through an external service provider, and the resulting nucleotide sequences were compared with reference sequences available in the National Center for Biotechnology Information (NCBI) database using the BLAST algorithm to determine species-level identity.
Quantitative Determination of BAs by HPLC
Only Enterococcus isolates that exhibited positive decarboxylase activity during preliminary in vitro screening were selected for quantitative BA analysis. BA quantification was performed using HPLC following acid extraction and dansyl chloride derivatization, adapted from a previously validated method (Sang et al., 2020).
Briefly, selected enterococcal strains were initially cultured in TSB at 37°C for 24 h. Subsequently, cultures were transferred into fresh TSB supplemented with 0.25% (w/v) of individual amino acid precursors (histidine, lysine, tyrosine, and ornithine hydrochloride) and incubated at 37°C for an additional 48 h to promote BA synthesis. For extraction, 1 mL of each culture was combined with an equal volume of 5% (w/v) trichloroacetic acid (TCA) to precipitate proteins, followed by centrifugation at 4°C for 10 min. The resulting supernatants were subjected to derivatization by sequential addition of 50 μL of 2 mol/L sodium hydroxide, 300 μL of dansyl chloride solution (10 mg/mL), and 100 μL of saturated sodium bicarbonate. The reaction was terminated by adding 50 μL of 25% ammonia solution, and the mixture was incubated in the dark at 25°C for 30 min to ensure complete derivatization.
Calibration curves were constructed using external standard solutions of histamine, tyramine, putrescine, cadaverine, and tryptamine at ten concentration levels within the linear range specified in Table 1. Each calibration level was analyzed in triplicate. Linearity was evaluated based on regression equations and correlation coefficients (R2). Chromatographic analysis was performed using a Shimadzu LC-2030 HPLC system (Kyoto, Japan) equipped with a reversed-phase C18 -column (Agilent ZORBAX Eclipse XDB-C18, 4.6 × 250 mm, 5 μm). The column temperature was maintained at 30°C, and the injection volume was set to 20 µL. The mobile phase consisted of ultrapure water (solvent A) and acetonitrile (solvent B), delivered at a constant flow rate of 1.0 mL/min. Detection was carried out using a photodiode array (PDA) detector set at 254 nm. The gradient elution program was applied as previously described. Separation was achieved using a gradient elution program as follows: 65–70% B from 0 to 5 min, 70–100% B from 5 to 14 min, 100% B from 14 to 18 min, 100–65% B from 18 to 20 min, and re-equilibration at 65% B from 20 to 22 min. Method performance was evaluated using a fit-for-purpose validation approach. Analytical parameters including linearity (R2), limits of detection (LOD), limits of quantification (LOQ), and linear working range were experimentally determined and are presented in Table 2. LOD and LOQ values were calculated based on the standard deviation of the response and the slope of the calibration curve (LOD = 3.3σ/ slope; LOQ = 10σ/ slope) (Atasoy et al., 2025). All samples were analyzed under identical exper imental conditions to ensure consistency and comparability of results. Internal standards were not employed in accordance with the original validated method.
Table 2. Regression parameters of biogenic amine compounds determined by HPLC method.
| Wavelength (nm) | Regression equation y = m (x) + n |
Correlation coefficient (r) |
Linear rangea (mg L−1) |
LODb (mg L−1) |
LOQb (mg L−1) | |
|---|---|---|---|---|---|---|
| Tryptamine | 254 | y = 3.27 109 (x) – 2.15 10–6 | 0.9995 | 0.10–50 | 0.014 | 0.043 |
| Putrescine | 254 | y = 4.98 109 (x) + 1.08 10–6 | 0.9998 | 0.10–200 | 0.023 | 0.071 |
| Cadaverine | 254 | y = 3.15 107 (x) + 3.46 10–6 | 0.9994 | 0.10–50 | 0.008 | 0.023 |
| Histamine | 254 | y = 4.12 109 (x) + 1.72 10–6 | 0.9989 | 0.05–50 | 0.001 | 0.003 |
| Tyramine | 254 | y = 5.76 109 (x) – 9.85 10–6 | 0.9995 | 0.10–250 | 0.005 | 0.014 |
a10 calibration points were studied for linearity range (n = 10); bThree replicates were performed (n = 3). LOD (µg/mL) = 3.3(SD of the response/slope); LOQ (µg/mL) = 10 (SD of the response/slope).
Statistical and chemometric analysis
All experiments were performed in triplicate (n = 3), and results are expressed as mean ± standard deviation. Statistical analyses were conducted using Minitab statistical software (version 17, Minitab Inc., UK). Differences among BA concentrations produced by different Enterococcus strains were evaluated using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Differences were considered statistically significant at P < 0.05.
Pearson’s correlation analysis was performed to assess relationships between individual BAs and total BA content. For multivariate analysis, principal component analysis (PCA) and hierarchical cluster analysis (HCA) were applied to explore patterns and relationships among BA profiles. Prior to analysis, data were mean-centered and standardized (auto-scaled) to eliminate the influence of scale differences among variables. PCA was performed using the covariance matrix, and the first two principal components (PC1 and PC2) were used to visualize the distribution of strains and their associations with BA variables. The explained variance for each principal component was calculated, and only components with significant contributions were considered in the interpretation. Despite the relatively small dataset, PCA was applied as an exploratory tool to identify patterns and relationships among variables. HCA was conducted using Ward’s linkage method based on squared Euclidean distance to classify Enterococcus strains according to similarities in their BA production profiles.
Nucleotide sequence accession numbers
The nucleotide sequences of the 16S rRNA genes from 135 Enterococcus isolates of the present study were submitted and deposited to the GenBank. An accession number was given in Table 3.
Table 3. Biochemical characterization and phenotypic profiles of aminogenic Enterococcus isolates.
| Molecularly characterized strains | Gram staining | Catalase activity | Bile esculin hydrolysis | Growth at 6.5% NaCl | Growth at pH 9.6 | Growth at 10°C | Growth at 45°C | Accession numbers |
|---|---|---|---|---|---|---|---|---|
| E. faecalis BLK14 | + | – | + | + | + | + | + | PZ188510 |
| E. faecalis BLK19 | + | – | + | + | + | + | + | PZ188511 |
| E. faecalis BLK45 | + | – | + | + | + | + | + | PZ188512 |
| E. faecalis BLK49 | + | – | + | + | + | + | + | PZ188513 |
| E. faecalis BLK70 | + | – | + | + | + | + | + | PZ188514 |
| E. faecalis BLK80 | + | – | + | + | + | + | + | PZ188515 |
| E. faecium BLK18 | + | – | + | + | + | + | + | PZ188759 |
| E. faecium BLK30 | + | – | + | + | + | + | + | PZ188760 |
| E. faecium BLK42 | + | – | + | + | + | + | + | PZ188761 |
| E. faecium BLK77 | + | – | + | + | + | + | + | PZ188762 |
| E. lactis BLK21 | + | – | + | + | + | + | + | PZ191240 |
| E. lactis BLK25 | + | – | + | + | + | + | + | PZ191241 |
| E. lactis BLK39 | + | – | + | + | + | + | + | PZ191242 |
| E. durans BLK54 | + | – | + | + | + | + | + | PZ191243 |
+: Positive; –: Negative
Results and Discussion
Out of the 56 fresh anchovy samples analyzed, a total of 24 Enterococcus strains were successfully recovered, representing a prevalence rate of 42.86% (data not shown). All isolates were subjected to preliminary morphological and cultural characterization. The recovered strains demonstrated the ability to grow under alkaline conditions (pH 9.6), in the presence of 6.5% NaCl, and across a temperature range of 10–45°C. In addition, all isolates were confirmed to be Gram-positive, catalase-negative, and capable of hydrolyzing esculin. Screening for BA production revealed that 14 of the 24 Enterococcus isolates (58.33%) exhibited aminogenic activity (Table 3). These 14 BA-producing strains were identified at the species level by 16S rRNA gene sequence analysis (Figure 1) indicating that the isolates consisted of 6 E. faecalis (42.86%), 4 E. faecium (28.57%), 3 E. lactis (21.43%), and 1 E. durans (7.14%) strains. Isolates were assigned to species level based on a ≥97% sequence similarity threshold against the GenBank database. The corresponding accession numbers for each strain are detailed in Table 3. However, it should be noted that 16S rRNA gene sequencing may offer limited resolution for species-level differentiation within the Enterococcus genus and should be interpreted with caution. Although enterococci are widely recognized as part of the natural microbiota of fermented foods (Atasoy et al., 2025; Dilireba et al., 2025; Elkassas and Fawzy Elbahy, 2016; Shanab et al., 2025), their presence and functional behavior in fresh fish have received comparatively limited attention. The relatively high isolation frequency observed in this study (42.86%) confirms that Enterococcus spp. can persist in fresh anchovy, likely reflecting both environmental contamination and their notable tolerance to adverse conditions such as elevated salinity, alkaline pH, and broad temperature ranges (Kukułowicz et al., 2024).
Figure 1. Amplified 16S rRNA fragments of the strains. L. O’Gene ruler DNA marker; 1. Enterococcus faecalis ATCC 29212 -(pozitif kontrol); 2. BLK18; 3. BLK19; 4. BLK21; 5. BLK22; 6. BLK25; 7. BLK30; 8. BLK36; 9. BLK39; 10. BLK40; 11. BLK42; 12. BLK45; 13. BLK49; 14. BLK53; 15. BLK54; 16. BLK57; 17. BLK65; 18. BLK 66, 19.BLK 68; 20. BLK 75; 21. Negative Control (Pure water).
All statistical analyses were conducted based on triplicate measurements (n = 3), and results were considered reliable within the defined experimental conditions. Statistical analysis revealed significant differences (p < 0.05) among strains for most BAs, as indicated by different superscript letters in Table 4. As summarized in Table 4, Enterococcus isolates recovered from fresh anchovy samples exhibited the capacity to synthesize several BAs. Tryptamine was the most frequently detected (13 isolates), followed by tyramine (11), histamine (10), cadaverine (6), and -putrescine (5). The concentrations of these compounds ranged from not detected (ND) to 236.928 mg/L for tryptamine, 307.638 mg/L for tyramine, 257.939 mg/L for histamine, 165.835 mg/L for cadaverine, and 56.317 mg/L for putrescine. The total BA production by Enterococcus strains varied between 85.070 ± 0.452 and 493.547 ± 1.411 mg/L. At the species level, E. faecalis exhibited BA levels between 207.289 ± 1.028 and 485.237 ± 2.083 mg/L, while E. lactis showed comparatively lower amounts (181.179 ± 1.017 to 300.432 ± 1.247 mg/L). E. faecium isolates produced total BA concentrations ranging from 250.851 ± 1.995 to 493.547 ± 1.411 mg/L, whereas E. durans displayed the lowest recorded level at 85.070 ± 0.452 mg/L. The BA production profiles exhibited significant -inter-species variation among the Enterococcus isolates examined (p < 0.05). BA production patterns differed markedly among the Enterococcus species. E. faecalis isolates were characterized by a predominance of tyramine formation, accompanied by moderate levels of tryptamine and histamine, whereas cadaverine and putrescine were consistently detected at relatively low levels. In contrast, E. lactis exhibited a more heterogeneous profile, with substantial variability in tryptamine production, moderate tyramine synthesis, and comparatively limited histamine formation. Notably, this species demonstrated a broader capacity for cadaverine and putrescine accumulation compared to E. faecalis. E. faecium isolates showed a strong ability to produce tryptamine, along with considerable amounts of tyramine and histamine, while cadaverine and putrescine were produced in limited amounts. Conversely, E. durans exhibited a restricted BA profile, with no detectable production of tryptamine, tyramine, or putrescine, while only limited amounts of histamine and cadaverine were produced. The dominant BAs detected in fresh anchovy samples included tyramine from E. faecalis BLK49, histamine and tryptamine from E. faecium BLK18 and BLK30, -respectively, as well as cadaverine and putrescine -produced by E. lactis BLK39 and BLK21.
Table 4. Biogenic amine concentrations (mg/L) produced by strains isolated from fresh anchovy samples.
| No | Strains | HIS (mg/L) |
TYR (mg/L) |
PUT (mg/L) |
CAD (mg/L) |
TYP (mg/L) |
Total BAs (mg/L) |
|---|---|---|---|---|---|---|---|
| 1 | E. faecalis BLK14 | 202.254 ± 1.978b | 264.327 ± 2.056d | ND | ND | 2.180 ± 0.054i | 468.761 ± 2.041c |
| 2 | E. faecalis BLK19 | ND | 165.756 ± 1.032ı | 7.814 ± 0.095d | ND | 33.719 ± 0.411f | 207.289 ± 1.028j |
| 3 | E. faecalis BLK45 | 202.608 ± 2.001b | 275.608 ± 2.103c | ND | ND | 7.375 ± 0.089g | 485.237 ± 2.083b |
| 4 | E. faecalis BLK49 | 16.004 ± 0.078f | 307.638 ± 2.378a | ND | ND | 43.763 ± 0.583e | 367.405 ± 2.375d |
| 5 | E. faecalis BLK70 | 21.604 ± 0.317e | 227.878 ± 2.122f | ND | 4.741 ± 0.071d | 43.909 ± 0.612e | 298.132 ± 2.120g |
| 6 | E. faecalis BLK80 | ND | 107.765 ± 0.894i | 11.656 ± 0.065b | 14.703 ± 0.073c | 90.662 ± 0.997c | 224.786 ± 0.968i |
| 7 | E. faecium BLK18 | 257.939 ± 1.431a | 191.971 ± 1.354g | ND | ND | 43.637 ± 0.631e | 493.547 ± 1.411a |
| 8 | E. faecium BLK30 | 13.923 ± 0.118g | ND | ND | ND | 236.928 ± 1.996a | 250.851 ± 1.995ı |
| 9 | E. faecium BLK42 | ND | 280.268 ± 2.011b | ND | ND | 4.210 ± 0.052ı | 284.478 ± 2.011h |
| 10 | E. faecium BLK77 | 35.242 ± 0.548d | 257.939 ± 1.998e | 3.094 ± 0.065e | 4.058 ± 0.071e | 56.660 ± 0.569d | 356.993 ± 1.997e |
| 11 | E. lactis BLK21 | 16.367 ± 0.078f | 35.853 ± 0.378j | 56.317 ± 0.501a | 0.549 ± 0.009f | 191.346 ± 1.248b | 300.432 ± 1.247f |
| 12 | E. lactis BLK25 | 3.904 ± 0.062h | 184.531 ± 1.024h | ND | ND | 0.593 ± 0.016j | 189.028 ± 1.023k |
| 13 | E. lactis BLK39 | ND | ND | 10.623 ± 0.100c | 165.835 ± 1.017a | 4.721 ± 0.069h | 181.179 ± 1.017l |
| 14 | E. durans BLK54 | 66.527 ± 0.478c | ND | ND | 18.543 ± 0.308b | ND | 85.070 ± 0.452m |
HIS: histamine; CAD: cadaverine; PUT: putrescine; TYR: tyramine; TYP: tryptamine; ND: Not Determined; (n=3, all parameters are given with their standard deviation). Values are expressed as mean ± standard deviation (n = 3). Different superscript letters (a–m) within the same column indicate statistically significant differences among strains (P < 0.05), as determined by one-way ANOVA followed by Tukey’s post hoc test.
More than half of the isolates (58.33%) exhibited aminogenic activity, highlighting Enterococcus spp. as significant contributors to BAs formation in fresh anchovy. This proportion is comparable to, or higher than, those reported in previous studies on fish and fishery products, where enterococci have been identified as important BA producers alongside members of the genera Morganella, Photobacterium, and Lactobacillus (Pertiwi et al., 2020; Visciano et al., 2020; Yilmaz, 2024). Notably, species-level identification revealed a predominance of E. faecalis and E. faecium among BA-producing isolates, in agreement with earlier findings indicating that these species possess a higher genetic potential for amino acid decarboxylation (Aktop and Şanlıbaba, 2026; Atasoy et al., 2025; Shanab et al., 2025; Wiśniewski and Barbieri, 2025).
The quantitative data demonstrated marked interspecies and intraspecies variability in BA production profiles. Tyramine and histamine emerged as the dominant amines, reaching concentrations exceeding 300 mg/L in some strains. Tyramine production was particularly pronounced in E. faecalis, whereas E. faecium strains were associated with the highest histamine and tryptamine levels. This variability is widely regarded in the literatures (Ba et al., 2017; Barbieri et al., 2019; Kang et al., 2017; Park et al., 2020; Wiśniewski and Barbieri, 2025; Zhang et al., 2025) as arising from strain-specific differences in the presence, regulation, and expression of decarboxylase genes, in addition to variations in substrate availability and complex metabolic interactions. Compared with other enterococcal species, E. durans showed relatively weak BA–forming capacity, indicating a lower overall contribution to aminogenic risk; however, its documented potential to synthesize histamine and cadaverine warrants continued consideration.
The presence of high concentrations of histamine and tyramine poses a significant concern for food safety and consumer risk assessment. Histamine is the primary causative agent of scombroid poisoning (Del Rio et al., 2017; Ruiz-Capillas and Herrero, 2019; Zhernov et al., 2023), while excessive tyramine intake may trigger hypertensive crises in sensitive individuals (Sadighara et al., 2024). In the present study, the highest tyramine levels, exceeding 300 mg/L, were comparable to the upper concentrations previously documented for Enterococcus strains isolated from dairy products, meat, and fermented plant-based foods (Park et al., 2020; Barbieri et al., 2021). Recent genomic and phenotypic studies suggest that the tyrosine decarboxylase operon is widely distributed in E. faecalis and E. faecium, supporting the hypothesis that high tyramine production may represent a recurrent metabolic trait in these species (Ladero et al., 2021). The present study extends these observations to enterococci originating from fresh fish, indicating that seafood may also harbor strains with substantial tyraminogenic capacity. In this study, histamine levels produced by specific E. faecium strains exceeded 250 mg/L in the screening medium. This high aminogenic potential suggests that enterococci could significantly contribute to histamine accumulation in fish products, particularly when subjected to temperature abuse or prolonged storage. Recent reports increasingly recognize LAB, including entero cocci, as non-negligible histamine producers alongside classical Gram-negative histamine-forming bacteria (Lo Magro et al., 2020; Pawul-Gruba et al., 2025; Wiśniewski and Barbieri, 2025).
Tryptamine production was another notable feature of the investigated isolates, particularly among E. fae cium strains, with concentrations exceeding 200 mg/L. Compared to histamine and tyramine, tryptamine has received less regulatory and toxicological attention; however, recent studies highlight its vasoactive and neuroactive properties and its potential to exacerbate adverse reactions when co-occurring with other BAs (Barbieri et al., 2021; Ozogul et al., 2023). The relatively high tryptamine levels observed in this study exceed those commonly reported for LAB in fermented foods (Arranz et al., 2025; Yilmaz 2024), suggesting that fish-derived substrates may favor enhanced tryptophan decarboxylation in certain enterococcal strains.
Cadaverine and putrescine were generally produced at lower levels, although E. lactis isolates displayed considerable variability, particularly with respect to cadaverine formation. While these diamines are not subject to specific legal limits, their toxicological relevance lies in their ability to potentiate histamine toxicity by inhibiting intestinal amine oxidases (Sánchez-Pérez et al., 2022). Recent risk assessments emphasize that even moderate levels of cadaverine and putrescine may significantly increase the health risk associated with histamine intake, especially in seafood products where multiple amines frequently co-occur (Tabanelli 2020).
Evaluation of total BA production further revealed that E. faecium isolates exhibited the highest cumulative BA levels, followed by E. faecalis, whereas E. durans showed a comparatively limited aminogenic profile. Similar species-related trends have been reported in recent studies investigating enterococci from fermented and non-fer mented foods, reinforcing the view that E. faecium and E. faecalis are among the most metabolically adaptable enterococcal species in food ecosystems (Atasoy et al., 2025; Banicod et al., 2025; Zhang et al., 2025). Although no official regulatory thresholds exist for total BA con tent, cumulative concentrations approaching 500 mg/L, as observed here, may still be of concern due to synergistic toxic effects.
Pearson’s correlation analysis was conducted to assess the relationships among individual BA compounds. Statistically significant positive correlations were identified between total BAs and histamine (r = 0.768, p < 0.002), as well as between total BAs and tyramine (r = 0.656, p < 0.011). In addition, a statistically meaningful correlation was detected between tryptamine and putrescine (r = 0.522, p < 0.049). No statistically significant associations were found among the other analyzed variables.
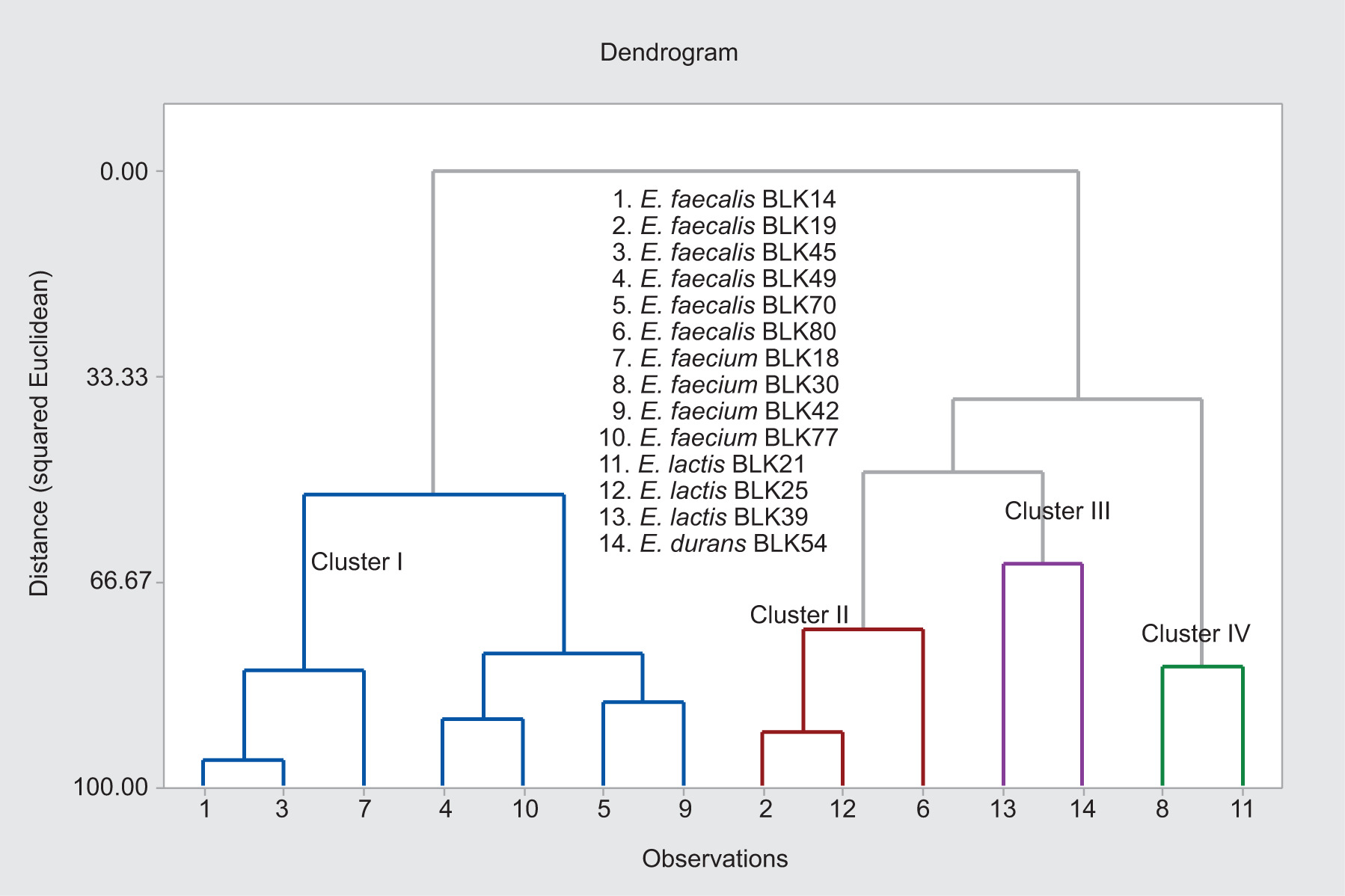
The HCA (Figure 2) revealed four principal clusters (Cluster I–IV), indicating distinct grouping patterns among Enterococcus strains based on their BA production profiles. Using this method, variables were classified into distinct clusters according to their degree of similarity, and the clustering structure was illustrated through a dendrogram. The results indicated that the 14 Enterococcus strains were organized into four principal, interrelated clusters. Cluster 1 comprised seven strains (BLK14, BLK45, BLK49, BLK70, BLK18, BLK42, and BLK77) and was primarily associated with tyramine and total BA levels. The overall connectivity among the binary groupings within this cluster was calculated as 52.63%. Cluster 2 comprised three components (strains BLK19, BLK80, and BLK25) associated with tyramine, total BAs, and tryptamine. These strains exhibited a strong degree of relatedness, indicating shared characteristics. The overall connectivity among the binary groupings within this cluster was determined to be 74.29%. Cluster 3 included two components (strains BLK39 and BLK54) associated with tryptamine and total BAs, whereas Cluster 4 comprised two components (strains BLK30 and BLK21) linked to cadaverine and total BA content. The highest similarity (95.76%) was detected between strains BLK14 and BLK45 within the tyramine–total BA profile, while a comparatively lower similarity (63.76%) was observed between strains BLK39 and BLK54 for the same variables. The overall relational strength among the seven components in Cluster 1 was calculated as 88.01%, whereas it was 47.14% for the three components in Cluster 2, 47.42% for the two components in Cluster 3, and 88.13% for the two components in Cluster 4. Among the least similar variable pairs, strains BLK19 and BLK308 associated with tyramine and total BAs showed a similarity of 37.05%. Furthermore, no statistically significant association was detected between strains BLK14 and BLK19. Overall, the clustering results highlight pronounced strain-dependent differences in BA production patterns, underscoring the heterogeneity of metabolic behaviors and emphasizing the necessity of strain-level evaluation for accurate assessment and management of BA-related risks in food systems.
Figure 2. Hierarchical cluster analysis (HCA) dendrogram showing the relationships among Enterococcus strains based on their biogenic amine production profiles. Clustering was performed using Ward’s linkage method and squared Euclidean -distance after data standardization. The dendrogram reveals four principal clusters (Cluster I–IV), grouping strains with similar biogenic amine production patterns. Species names and corresponding strain codes are indicated for each isolate.
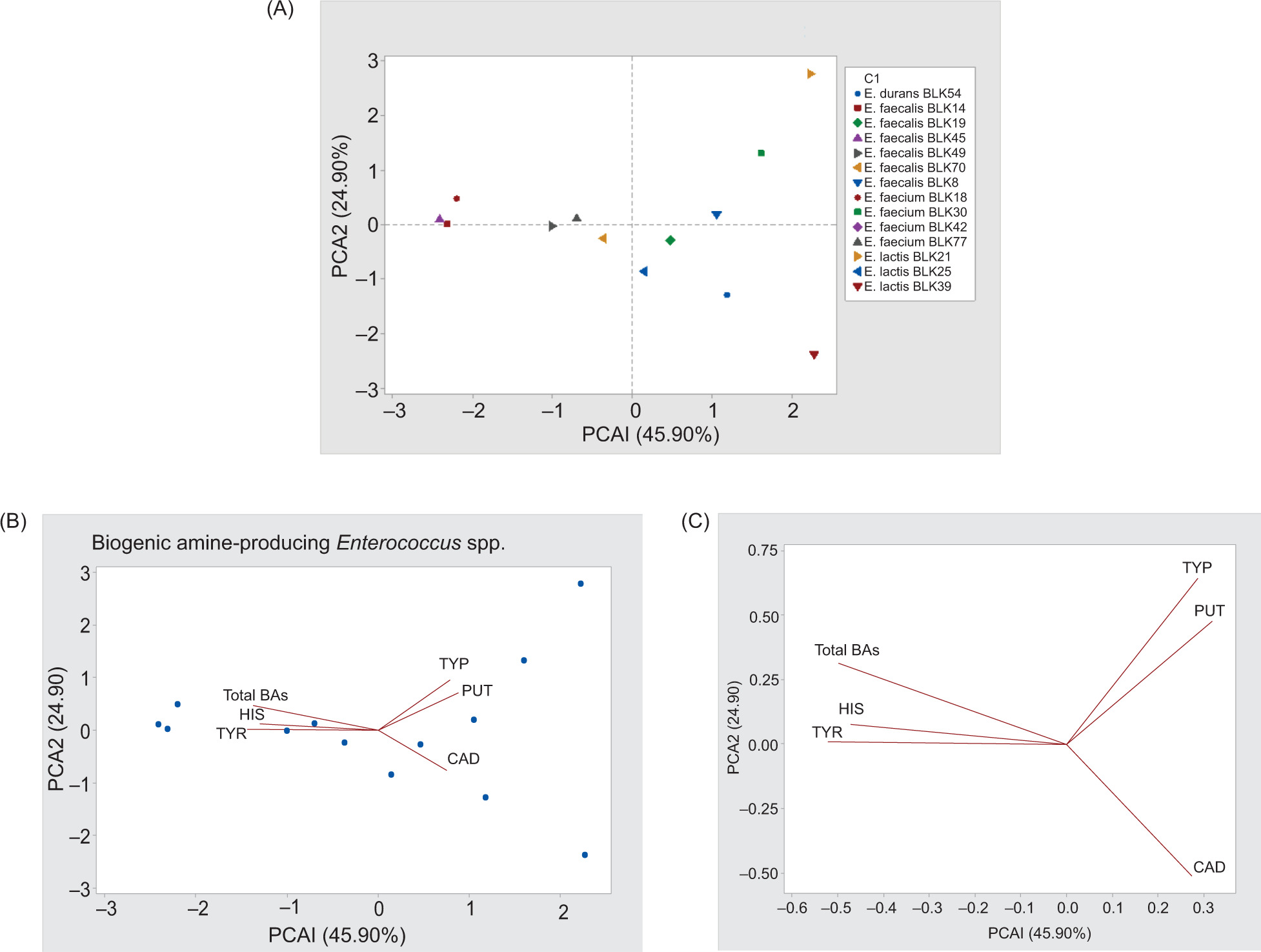
To obtain a more integrated insight into the patterns and interrelationships among the variables associated with BA composition in fresh anchovy, PCA was applied (Ghayoomi et al., 2024). The data were mean-centered and standardized prior to analysis to minimize the influence of scale differences among variables. The first two principal components (PC1 and PC2) explained 45.90% and 24.90% of the total variance (70.80%), respectively, indicating that a substantial proportion of the variability in the dataset could be captured in a -two-dimensional space. The PCA score plot revealed a clear separation of strains according to their BA production patterns. Strains located in the same quadrant exhibited similar profiles, whereas those positioned in opposite directions were characterized by distinct BA compositions. Loadings indicated that specific BAs contributed differently to the observed variation. For instance, histamine and tyramine were strongly associated with strains positioned along the positive axis of PC1, while other amines contributed to separation along PC2. Overall, PCA provided a useful multivariate approach to visualize clustering patterns and relationships among strains, supporting the variability observed in individual BA measurements. The PC1, explaining 45.90% of the total variance, showed positively associated with tyramine, tryptamine, and putrescine, while exhibiting a negative association with cadaverine (Figure 3B, 3C; regions 2 and 4). The PC2 accounted for 24.90% of the overall variance and was positively correlated with total BAs, histamine, and tyramine (Figure 3B, C; region 1).
Figure 3. Principal component analysis (PCA) score plot showing the distribution of Enterococcus strains based on their biogenic amine production profiles. Data were mean-centered and auto-scaled prior to analysis. The first two principal components (PC1 and PC2) explained 45.90% and 24.90% of the total variance (70.80%), respectively. Strains with similar biogenic amine profiles are grouped closely, while the direction and length of vectors indicate the contribution of individual biogenic amines to the overall variability. (A) Principal component analysis (PCA) score plot, and (B–C) loading plots for biogenic amines in fresh anchovy.
A combined analysis of Figures 3A and 3C indicated that five strains located in region 1 (E. faecium BLK18, E. faecalis BLK45, E. faecalis BLK14, E. faecalis BLK49, and E. faecium BLK77) were positively associated with total BAs, histamine, and tyramine. In contrast, two strains positioned in region 3 (E. faecalis BLK70 and E. faecium BLK42) showed negative loadings for tyramine. The close overlap of these strains in Figure 3A and B suggests that they produced highly comparable BA levels. Furthermore, strains assigned to region 4 (E. faecalis BLK19, E. lactis BLK25, E. durans BLK54, and E. lactis BLK39) exhibited negative correlations with cadaverine, whereas three strains clustered in region 2 (E. faecalis BLK80, E. faecium BLK30, and E. lactis BLK21) were positively associated with tryptamine and putrescine. Consistent with recent findings demonstrating significant strain-dependent variability in BA production among Enterococcus isolates from fermented foods (Wiśniewski and Barbieri, 2025), which report high inter-strain differences in tyramine and other BA levels, our PCA results confirm that BA profiles in fresh anchovy are not uniform across strains, underscoring the importance of strain-level characterization for accurate food safety assessment and control strategies
The present results underline that Enterococcus spp. -isolated from fresh anchovy should be regarded as potential contributors to BA accumulation rather than harmless background microbiota. The detection of strains capable of producing histamine and tyramine at levels close to or exceeding regulatory limits highlights the importance of incorporating enterococci into seafood safety monitoring strategies. These findings support recent calls for a broader, risk-based approach to BA control in fish that considers both classical spoilage bacteria and aminogenic LAB (Ozogul et al., 2023). The innovative aspect of this study lies in the comprehensive, species-level characterization of BA-producing Enterococcus strains isolated specifically from fresh anchovy, coupled with quantitative profiling of multiple BAs. To the best of our knowledge, data address ing the aminogenic potential of E. lactis and E. durans in fresh marine fish are scarce. By addressing this gap, the present findings broaden the current understanding of BA-related hazards in seafood and support the inclusion of enterococci in routine microbiological risk assessments of fresh fish. Accordingly, monitoring programs should extend beyond classical histamine-producing bacteria to encompass Enterococcus species with demonstrated aminogenic capacity. Overall, the results suggest that monitoring strategies for fresh anchovy and similar fish species should not be limited to traditional histamine-producing bacteria but should also encompass enterococci with proven aminogenic capacity. Further studies integrating genomic analysis of decarboxylase genes and evaluating BA formation under realistic storage conditions would be valuable to better assess the actual risk posed by these microorganisms along the seafood supply chain.
It should be noted that the characterization of BA-producing isolates in this study primarily focused on phenotypic screening. While phenotypic methods provide essential data on the functional activity and metabolic output of microorganisms, the absence of molecular analysis of specific amino acid decarboxylase genes limits the current study’s defined scope. Future research incorporating genotypic characterization and gene expression analysis will be instrumental in providing a deeper understanding of the genetic potential and regulatory mechanisms governing BA production in these strains.
Conclusion
This study demonstrates that fresh anchovy (Engraulis encrasicolus) can harbor Enterococcus species with a pronounced capacity to produce BAs, underscoring their relevance as previously underrecognized contributors to chemical food safety hazards in non-fermented fish products. The predominance of E. faecalis and E. faecium, together with their strain-dependent and -species-specific aminogenic profiles, highlights the complexity of BA formation and the limitations of relying solely on traditional histamine-producing bacteria in seafood safety assessments. The detection of elevated concentrations of histamine, tyramine, and other BAs at levels approaching or exceeding those associated with adverse health effects emphasizes the need for a risk-based perspective that considers cumulative and synergistic toxicity. Within a One Health framework, the presence of aminogenic Enterococcus spp. in fresh fish represents an intersection of environmental contamination, food processing practices, and human exposure, reinforcing the importance of integrated surveillance across the seafood supply chain. Overall, the findings support the inclusion of Enterococcus spp. in routine monitoring and risk assessment strategies for fresh fish products, particularly in regions where anchovy constitutes a major dietary component. Future research combining genomic characterization of aminogenic determinants with predictive modeling under realistic storage conditions will be essential to refine risk management approaches and inform evidence-based regulatory decisions on BAs beyond -histamine alone.
Availability of Data and Materials
The nucleotide sequences of the 16S rRNA genes from Enterococcus isolates analyzed in the present study have been submitted to and archived in GenBank.
Mandatory Disclosure on Use of Artificial Intelligence
The authors didn’t use Artificial Intelligence for language editing, clarity improvement, or any other function for paper improvement.
Authors’ Contributions
All authors contributed equally to the conception and design of the study, data acquisition, analysis and interpretation of the results, and the drafting and critical revision of the manuscript. All authors approved the final version of the manuscript.
Conflicts of Interests
The authors declare that they have no competing interests.
Funding
None.
REFERENCES
Akpomie, O.O., Ejechi, B.O., Banach, A.M., Adewuyi, I., Ayobola, E.D., Akpomie, K.G., Ghosh, S., and Ahmadi, S. 2022. Biogenic amine production from processed animal and plant protein-based foods contaminated with Escherichia coli and Enterococcus feacalis. Journal of Food Science and Technology 59(12): 4880–4888. 10.1007/s13197-022-05576-0
Aktop, S., and Şanlıbaba, P. 2026. Isolation and identification of biogenic amines producer Enterococcus strains isolated from traditional cheeses. Italian Journal of Food Science 38(1): 47–59. 10.15586/ijfs.v38i1.3302
Al Bulushi, I., Poole, S., Deeth, H.C., and Dykes, G.A. 2009. Biogenic amines in fish: roles in intoxication, spoilage, and nitrosamine formation--a review. Critical Reviews in Food Science and Nutrition 49(4): 369–377. 10.1080/1040839080206751460
Arranz, D., Fernández, E., Szekeres, B., Carvalho, A., Rio, B.D., Redruello, B., and Alvarez, M.A. 2025. Tryptamine accumulates in cheese mainly via the decarboxylation of tryptophan by lactic acid bacteria. Food Research International 199: 115380. 10.1016/j.foodres.2024.115380
Arulkumar, A., Paramithiotis, S., and Paramasivam, S. 2023. Biogenic amines in fresh fish and fishery products and emerging control. Aquaculture and Fisheries 8(4): 431–450. 10.1016/j.aaf.2021.02.001
Atasoy, G., Şanlıbaba, P., Vural, N., and Anlı, R.E. 2025. Production of biogenic amines by Enterococcus strains from green and black table olives in Türkiye. Italian Journal of Food Science 37(3): 310–324. 10.15586/ijfs.v37i3.3043
Ba, H.V., Seo, H.W., Cho, S.H., Kim, Y.S., Kim, J.H., Park, B.Y., Kim, H.W., Ham, J.S., and Seong, P.N. 2017. Utilisation possibility of Enterococcus faecalis isolates from neonate’s faeces for production of fermented sausages as starter cultures. International Journal of Food Science & Technology 52(7): 1660–1669, 10.1111/ijfs.13440
Banicod, R.J.S., Ntege, W., Njiru, M.N., Abubakar, W.H., Kanthenga, H.T., Javaid, A., and Khan, F. 2025. Production and transformation of biogenic amines in different food products by the metabolic activity of the lactic acid bacteria. International Journal of Food Microbiology. 428: 110996. 10.1016/j.ijfoodmicro.2024.110996
Barbieri, F., Montanari, C., Gardini, F., and Tabanelli, G. 2019. Biogenic amine production by lactic acid bacteria: A review. Foods 8(1): 17. 10.3390/foods8010017
Barbieri, F., Tabanelli, G., Montanari, C., Dall’Osso, N., Šimat, V., Smole Možina, S., Baños, A., Ozogul, F., Bassi, D., Fontana, C., and Gardini, F. 2021. Mediterranean spontaneously fermented sausages: Spotlight on microbiological and quality features to exploit their bacterial biodiversity. Foods 10: 2691. 10.3390/foods10112691
Bayesen, D., and Yüksel, Z. 2023. Biyojen Aminlerin Fermente Gıdalarda Oluşum Süreci ve Etkileri. İstanbul Rumeli Üniversitesi Sağlık Bilimleri Dergisi 2(2): 50–65.
Beasley, S.S., and Saris, P.E. 2004. Nisin-producing Lactococcus lactis strains isolated from human milk. Applied and Environmental Microbiology 70(8): 5051–5053. 10.1128/AEM.70.8.5051-5053.2004
Bover-Cid, S., and Holzapfel, W.H. 1999. Improved screening procedure for biogenic amine production by lactic acid bacteria. International Journal of Food Microbiology 53(1): 33–41. 10.1016/S0168-1605(99)00152-X
Buňka, F., Budinský, P., Zimáková, B., Merhaut, M., Flasarová, R., Pachlová, V., Kubáň, V., and Buňková, L. 2013. Biogenic amines occurrence in fish meat sampled from restaurants in region of Czech Republic. Food Control 31 (1): 49–52. 10.1016/j.foodcont.2012.09.044
Connil, N., Le Breton, Y., Dousset, X., Auffray, Y., Rincé, A., and Prévost, H. 2002. Identification of the Enterococcus faecalis tyrosine decarboxylase operon involved in tyramine production. Applied and Environmental Microbiology 68(7): 3537–3544. 10.1128/AEM.68.7.3537-3544.2002
Çakmak, T., and Sancak, Y.C. 2023. Determination of biogenic amine formation, microbiological and sensory changes in Carp (Cyprinus carpio L., 1758) stored at cold (4°C). Van Veterinary Journal 34(1): 32–42. 10.36483/vanvetj.1209788
Dankar, I., Melhem, A., Serhan, M., and Hassan, H.F. 2025. Microbial and processing factors affecting biogenic amine formation and accumulation in dairy: a narrative review. Applied Food Research 5(2): 101376. 10.1016/j.afres.2025.101376
Dara, A., Akhondzadeh Basti, A., Mahasti Shotorbani, P., Tamadoni Jahromi, S., Jabar Zadeh Shiadeh, M., and Azizian, A. 2025. Investigating histamine levels, microbial and chemical properties in industrial and traditional drying methods of anchovy fish in Qeshm Island. Archives of Razi Institute 80(1): 131–137. 10.32592/ARI.2025.80.1.131
Del Rio, B., Redruello, B., Linares, D.M., Ladero, V., Fernandez, M., Martin, M.C., Ruas-Madiedo, P., and Alvarez, M.A. 2017. The dietary biogenic amines tyramine and histamine show synergistic toxicity towards intestinal cells in culture. Food Chemistry 218: 249–255. 10.1016/j.foodchem.2016.09.046
Dilireba, E., Lu, Q., Chang, L., and Luo, F. 2025. Antibiotic resistance profile and molecular mechanism of enterococci in traditional fermented yogurts. Journal of Dairy Science 109 (2): 1006-1019. 10.3168/jds.2025-27212
Ding, T., and Li, Y. 2024. Biogenic amines are important indices for characterizing the freshness and hygienic quality of aquatic products: A review. LWT. 194: 115793. 10.1016/j.lwt.2024.115793
EFSA (European Food Safety Authority). 2011. Scientific opinion on risk based control of biogenic amine formation in fermented foods. Panel on Biological Hazards (BIOHAZ). Efsa J. 9(10): 2393–2487. 10.2903/j.efsa.2011.2393
Elkassas, W., and Fawzy Elbahy, E. 2016. Determination of some-biogenic amines levels in some cheese varieties. Assiut Veterinary Medical Journal 62(148): 84–91. 10.21608/avmj.2016.169220
Fadhlaoui-Zid, K., Curiel, J.A., Landeta, G., Fattouch, S., Reverón, I., De Las Rivas, B., Sadok, S., and Muñoz, R. 2012. Biogenic amine production by bacteria isolated from ice-preserved sardine and mackerel. Food Control 25(1): 89–95.
Gao, X., Li, C., He, R., Zhang, Y., Wang, B., Zhang, Z.H., and Ho, C.T. 2023. Research advances on biogenic amines in traditional fermented foods: Emphasis on formation mechanism, detection and control methods. Food Chemistry. 405, Part A: 134911. 10.1016/j.foodchem.2022.134911
Goa, T., Beyene, G., Mekonnen, M., and Gorems, K. 2022. Isolation and characterization of lactic acid bacteria from fermented milk produced in Jimma Town, Southwest Ethiopia, and evaluation of their antimicrobial activity against selected pathogenic bacteria. International Journal of Nutrition and Food Sciences. 2022: 2076021. 10.1155/2022/2076021
Ghayoomi, H., Edalatian Dovom, M.R., Habibi Najafi, M.B., and Pourfarzad, A. 2024. The principal component analysis of key and significant features of the safety and nutritional value of Mahyaveh sauce. Food Science and Nutrition 12(4): 2896–2907. 10.1002/fsn3.3970
In, J.J., Shim, K.B., Lee, J.B., Bae, Y.J., Kwon, G.Y., Lee, H.R., Park, S., and Sohn, S.K. 2024. Effect of activated carbon-based two-stage adsorption on biogenic amine reduction and quality of anchovy fish sauce at industrial scale. Food Chemistry. 458: 140169. 10.1016/j.foodchem.2024.140169
Javaid, A., Tabassum, N., Karthikeyan, A., Kim, Y.M., Jung, W.K., and Khan, F. 2025. Prevalence and stable acquisition of biogenic amine-synthesizing genes in lactic acid bacteria across diverse niches: implications for food safety and human health. NPJ Science Food 9(1): 268. 10.1038/s41538-025-00667-6
Kang, H.R., Kim, H.S., Mah, J.H., Kim, Y.W., and Hwang, H.J. 2017. Tyramine reduction by tyrosine decarboxylase inhibitor in Enterococcus faecium for tyramine controlled cheonggukjang. Food Science and Biotechnology 27(1): 87–93. 10.1007/s10068-017-0205-0
Kočar, D., Köse, S., Tufan, B., Ščavničar, A., and Pompe, M. 2021. Determination of biogenic amines in fresh fish and processed fish products using IC-MS/MS. Foods 10(8): 1746. 10.3390/foods10081746
Kukułowicz, A., Steinka, I., and Gardocka, M. 2024. Enterococcus spp. in fish: Analysis of the presence and resistance in samples from Tri-City, Poland. PLoS One 19(7): e0306826. 10.1371/journal.pone.0306826
Ladero, V., Calles-Enríquez, M., Fernández, M., and Alvarez, M.A. 2021. Toxicological effects of dietary biogenic amines. Current Nutrition & Food Science 6(2): 145–156. 10.2174/157340110791233256
Li, P., Chen, Y., Bai, J., Yang, H., He, P., and Zeng, J. 2025. The-determination of eight biogenic amines using MSPE-UHPLC-MS/MS and their application in regard to changes in these biogenic amines in traditional chinese dish-pickled swimming crabs. Molecules 30(6): 1353. 10.3390/molecules30061353
Lo Magro, S., Summa, S., Iammarino, M., D’Antini, P., Marchesani, G., Chiaravalle, A.E., and Muscarella, M. 2020. A 5-Years (2015–2019) Control Activity of an EU Laboratory: Contamination of histamine in fish products and exposure assessment. Applied Sciences. 10: 8693. 10.3390/app10238693
Ma, X., Bi, J., Li, X., Zhang, G., Hao, H., and Hou, H. 2021. Contribution of microorganisms to biogenic amine accumulation during fish sauce fermentation and screening of novel starters. Foods 10(11): 2572. 10.3390/foods10112572
Maddaloni, L., Grasso, S., De Gara, L., Pennazza, G., Zompanti, A., Rapa, M., Ruggieri, R., Vinci, G., and Santonico, M. 2021. An electrochemical sensor for monitoring biogenic amines in anchovies as quality and safety index. Sensors & Actuators, B: Chemical. 347: 130648. 10.1016/j.snb.2021.130648
Meng, J., Yang, Q., Wan, W., Zhu, Q., Zeng, X. 2022. Physicochemical properties and adaptability of amine-producing Enterobacteriaceae isolated from traditional Chinese fermented fish (Suan yu). Food Chemistry. 369: 130885. 10.1016/j.foodchem.2021.130885
Muhtar, A., Jian, P., Muhammad, T., Zhao, J., Dolkun, A., Zhou, T., and Piletsky, S.A. 2025. A dummy template molecularly imprinted polymer-coated fiber array extraction for simultaneous HPLC analysis of eight biogenic amines in fermented horse milk. Analytica Chimica Acta. 1352: 343901. 10.1016/j.aca.2025.343901
Mutalipassi, M., D’Anza, E., Pugliano, M., Firmamento, R., Murano, C., Ruocco, N., Pennesi, C., Procaccini, G., Romeo, T., Terlizzi, A., and Peretti, V. 2024. Casting light on the European anchovy: from biology to conservation and industry. Frontiers in Ecology and Evolution. 12: 1352055. 10.3389/fevo.2024.1352055
Ozogul, F., Durmuş, M., Kosker, A.R., Özkütük, A.S., Kuley, E., Yazgan, H., Yazgan, R., Simat, V., and Ozogul, Y. 2023. The impact of marine and terrestrial based extracts on the freshness quality of modified atmosphere packed sea bass fillets. Food Biosciences. 53: 102545. 10.1016/j.fbio.2023.102545
Özyurt, G., Özkütük, A.S., and Boğa, E.K. 2022. Türk Marketlerinde Satışa Sunulan Çeşitli Balık Ürünlerinin Biyojenik Amin ve Trimetilamin İçerikleri. Acta Aquatica Turcica 18(1): 13–23. 10.22392/actaquatr.906996
Park, Y.K., Jin, Y.H., Lee, J.-H., Byun, B.Y., Lee, J., Jeong, K.C., and Mah, J.-H. 2020. The role of Enterococcus faecium as a key producer and fermentation condition as an ınfluencing factor in tyramine accumulation in Cheonggukjang. Foods 9(7): 915. 10.3390/foods9070915
Pawul-Gruba, M., Denis, E., Kiljanek, T., and Osek, J. 2025. Prevalence of biogenic amines and their relation to the bacterial content in ripened cheeses on the retail market in Poland. Foods. 14: 2478. 10.3390/foods14142478
Pertiwi, R.M., Nurilmala, M., Abdullah, A., Nurjanah, Yusfiandayani, R., and Sondita, M.F.A. 2020. Deteksi bakteri pembentuk amina biogenik pada ikan Scombridae secara multiplex PCR. Jurnal Pengolahan Hasil Perikanan Indonesia 23(2): 359–371.
Pons-Sánchez-Cascado, S., Bover-Cid, S., Veciana-Nogués, M.T., and Vidal-Carou, M.C. 2005. Amino acid-decarboxylase activity of bacteria isolated from ice-preserved anchovies. European Food Research and Technology. 220: 312–315. 10.1007/s00217-004-1095-y
Ruiz-Capillas, C., and Herrero, A.M. 2019. Impact of biogenic amines on food quality and safety. Foods 8(2): 62. 10.3390/foods8020062
Sadighara, P., Bekheir, S.A., Shafaroodi, H., Basaran, B., and Sadighara, M. 2024. Tyramine, a biogenic agent in cheese: amount and factors affecting its formation, a systematic review. Food Production, Processing and Nutrition 6: 30. 10.1186/s43014-024-00223-x
Sánchez-Pérez, S., Comas-Basté, O., Costa-Catala, J., Iduriaga-Platero, I., Veciana-Nogués, M.T., Vidal-Carou, M.C., and Latorre-Moratalla, M.L. 2022. The rate of histamine degradation by diamine oxidase ıs compromised by other biogenic amines. Frontiers in Nutrition 9: 897028. 10.3389/fnut.2022.897028
Sang, X., Li, K., Zhu, Y., Ma, X., Hao, H., Bi, J., and Hou, H. 2020. The impact of microbial diversity on biogenic amines formation in grasshopper sub shrimp paste during the fermentation. Frontiers in Microbiology 11: 782. 10.3389/fmicb.2020.00782
Shanab, O., Fareed, F., Nassar, A.Y., Abd-Elhafeez, H.H., Ahmed, A.S., and El-Zamkan, M. 2025. Molecular characterization of histidine and tyrosine decarboxylating Enterococcus species isolated from some milk products. BMC Microbiology 25: 234. 10.1186/s12866-025-03940-6
Sivamaruthi, B.S., Kesika, P., and Chaiyasut, C. 2021. A narrative review on biogenic amines in fermented fish and meat products. Journal of Food Science and Technology 58(5): 1623–1639. 10.1007/s13197-020-04686-x
Tabanelli, G. 2020. Biogenic amines and food quality: Emerging challenges and public health concerns. Foods 9: 859. 10.3390/foods9070859
Ucar, Y., and Ozogul, F. 2024. Biogenic amines in seafood. Food Bulletin. 3(1): 9–15. 10.61326/foodb.v3i1.258
Visciano, P., Schirone, M., Tofalo, R., and Suzzi, G. 2012. Biogenic amines in raw and processed seafood. Frontiers in Microbiology. 3: 188. 10.3389/fmicb.2012.00188
Visciano, P., Schirone, M., and Paparella, A. 2020. An overview of histamine and other biogenic amines in fish and fish products. Foods 9(12): 1795. 10.3390/foods9121795
Wiśniewski, P., and Barbieri, F. 2025. Molecular ıdentification and biogenic amine production capacity of Enterococcus faecalis strains ısolated from raw milk. International Journal of Molecular Sciences. 26(21): 10480. 10.3390/ijms262110480
Yilmaz N. 2024. Quantitative analysis of biogenic amine production of different lactic acid bacteria isolated from ready-to-eat packaged fish products. Veterinary Research Forum. 15(10): 537–543. 10.30466/vrf.2024.2024103.4193
Zeng, X., Xia, W., Yang, F., ad=nd Jiang, Q. 2013. Changes of-biogenic amines in Chinese low-salt fermented fish pieces (Suan yu) inoculated with mixed starter cultures. International Journal of Food Science & Technology 48 (4): 685–692, 10.1111/ijfs.12010
Zhang, Y., Shan, B., Gong, J., and Hu, Y. 2022. Mechanism of biogenic amine synthesis of Enterococcus faecium isolated from Sanchun ham. Food Science & Nutrition. 10: 2036–2049. 10.1002/fsn3.2820
Zhang, X., Chi, H., Peng, D., Jiang, M., Wang, C., Zhang, H., Kang, W., and Li, L. 2025. Integrated metagenomic and LC–MS/MS analysis reveals the biogenic amine-producing strains of two typical chinese traditional fish products: Fermented mandarin fish (Siniperca chuatsi) and semi-dried yellow croaker (Larimichthys crocea). Foods 14(6): 1016. 10.3390/foods14061016
Zhernov, Y.V., Simanduyev, M.Y., Zaostrovtseva, O.K., Semeniako, E.E., Kolykhalova, K.I., Fadeeva, I.A., Kashutina, M.I., Vysochanskaya, S.O., Belova, E.V., Shcherbakov, D.V., Sukhov, V.A., Sidorova, E.A., and Mitrokhin, O.V. 2023. Molecular mechanisms of scombroid food poisoning. International Journal of Molecular Sciences 24(1): 809. 10.3390/ijms24010809