
Download
ORIGINAL ARTICLE
Production of biogenic amines by Enterococcus strains from green and black table olives in Türkiye
Gülsüm Atasoy1, Pınar Şanlıbaba1, Nilüfer Vural2, Rahmi Ertan Anlı1*
1Department of Food Engineering, Faculty of Engineering, Ankara University, Ankara, Türkiye;
2Biotherapeutic Products Research and Development Program, Department of Traditional, Complementary and Integrative Medicine, Institute of Public Health, Ankara Yıldırım Beyazit University, Ankara, Türkiye
Abstract
Table olives are among the most significant traditional fermented vegetables in Türkiye, with their global consumption steadily increasing. This study aimed to investigate the presence of biogenic amine (BA)-producing Enterococcus strains in traditional table olives. A total of 186 probable enterococcal isolates were identified from 460 table olive samples, including 240 green and 220 black olives. The ability of Enterococcus spp. to produce five BAs, including tyramine, cadaverine, putrescine, tryptamine, and histamine, was evaluated. The decarboxylase activity of Enterococcus isolates was analyzed using a modified decarboxylase medium. Among these, 71 isolates were determined as BA producers. Species-level identification through 16S rDNA sequence analysis classified these strains as E. faecium (20 isolates), E. faecalis (31 isolates), and E. lactis (20 isolates). Concentrations of BAs were quantified through high-performance liquid chromatography. The maximum concentrations of tyramine, cadaverine, putrescine, tryptamine, and histamine detected in the samples were 257.939 mg/L, 13.923 mg/L, 139.620 mg/L, 30.562 mg/L, and 7.985 mg/L, respectively. The total content of BAs produced by Enterococcus strains from green olives varied between 1.018 mg/L and 259.324 mg/L, while those from black olives ranged from 1.831 mg/L and 214.678 mg/L. Predominant BA detected in green olives was tyramine (257.939 mg/L). Similarly, in black olives, the highest BA levels were recorded for tyramine (207.618 mg/L). These findings highlight the significant presence of BA-producing Enterococcus strains in table olives, emphasizing the need for monitoring and control strategies to ensure food safety.
Key words: biogenic amine, food safety, Enterococcus spp, table olive
*Corresponding Author: Rahmi Ertan Anlı, Department of Food Engineering, Faculty of Engineering, Ankara University, Ankara 06830, Türkiye. Email: [email protected]
Academic Editor: Prof. Mariella Calasso — (SIMTREA) – University of Bari, Italy
Received: 27 February 2025; Accepted: 24 April 2025; Published: 1 July 2025
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Biogenic amines (BAs) are small nitrogen-containing molecules formed in living organisms through the decarboxylation of L-amino acids or their derivatives during the fermentation of food (Ghorbani et al., 2021; Guba et al., 2022; Moniente et al., 2022). BAs in fermented food products are generated due to uncontrolled microbial enzymatic activity of specific microorganisms, particularly those capable of producing amino acid decarboxylases (Gao et al., 2022; Ovalle-Marmolejo et al., 2023). Enzymes such as histidine decarboxylase or lysine decarboxylase transform amino acids such as histidine and lysine into histamine and cadaverine, respectively (Barbieri et al., 2019). These enzymes can be endogenous, originating from raw ingredients, or exogenous, produced by microbes during fermentation (Ahangari et al., 2021; Müller et al., 2022). The process not only aids in pH regulation but also provides an adaptive mechanism against acid stress (Barbieri et al., 2019; Pereira et al., 2009). The energy dynamics involved in this transformation also contribute to microbial survival (Li and Lu, 2020).
The type and quantity of BAs formed vary significantly based on the nature of the food and the microorganisms present (Lázaro et al., 2015). In fermented products, the primary microbial groups associated with BAs’ production are typically certain lactic acid bacteria (LAB), such as Enterococcus, Lactococcus, Lactobacillus, Carnobacterium, Leuconostoc, and Pediococcus strains (Ghorbani et al., 2021; Lázaro et al., 2015). Many carboxylase-positive species within these groups can concurrently produce various BAs. LAB are generally considered nutritionally beneficial, and the production of BAs has been linked to a protective effect against the acidic environment commonly found in fermented foods (Ovalle-Marmolejo et al., 2023).
Biogenic amines, characterized as heat-stable, nonvolatile organic bases with a pH of >9, have been detected in various protein- and amino acid-rich fermented foods, including fermented vegetables, sausages, cheese, beer, and wine (Huang et al., 2021; Kim et al., 2022; Li et al., 2022; Luo et al., 2022; Molaei et al., 2019). BAs act as signaling molecules in the body by contributing to various metabolic processes, such as hormone and alkaloid synthesis and heart protection (Ovalle-Marmolejo et al., 2023); however, when their breakdown is impaired or enzyme systems are overwhelmed, they may become toxic—particularly as tyramine, histamine, and β-phenylethylamine cause inflammatory reactions, and cadaverine and putrescine may worsen these effects by blocking histamine degradation (Luo et al., 2022; Mah et al., 2019; Ovalle-Marmolejo et al., 2023).
Several national and international regulatory bodies, including Health Canada, the US Food and Drug Administration (US FDA), and the European Food Safety Authority (EFSA), have established action thresholds for histamine concentrations, specifically in fish and fish-derived products, because of their high susceptibility to histamine accumulation (Turna et al., 2024). However, standardized regulatory guidelines are currently lacking for other categories of fermented foods and other BAs, such as tyramine and β-phenylethylamine, despite their well-documented toxicological implications. In general, it is recommended that the total BA content in food products should not exceed 1,000 mg/kg, with specific limits proposed for individual amines, such as β-phenylethylamine (30 mg/kg), tyramine (100–800 mg/kg), and histamine (200 mg/kg) (Akpomie et al., 2022; Jeon et al., 2018; Mah et al., 2019). Furthermore, the EFSA, in collaboration with the Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO, 2013) Expert Committee on Food Safety, has defined the no observed adverse effect level (NOAEL) for histamine intake as 50 mg per meal, underscoring its toxicological significance and relevance to public health risk management (Banicod et al., 2025). Moreover, certain BAs, such as histamine and tyramine, serve as indicators of food spoilage, and the presence of exogenous amines in fermented foods diminishes the sensory character by imparting unpleasant aromas (Luo et al., 2022; Silva et al., 2020; Yilmaz et al., 2022). These toxic compounds are of critical concern as they have the potential to impact human health as well (Kalinowska and Tobiszewski, 2023; Shalaby et al., 2016).
The trade standard for table olives, as defined by the International Olive Oil Council (IOOC, 2004), characterizes table olives as the healthy fruit derived from specific cultivars of olive trees (Olea europaea L.). These cultivars are selected based on attributes, such as fruit size, shape, flesh-to-stone ratio, texture, taste, firmness, and the ease with which the flesh detaches from the stone, rendering them particularly suitable for processing. The trade standard specifies that table olives have treatments to eliminate their natural bitterness and are preserved either through natural fermentation or thermal processing, with or without the inclusion of preservatives, and are packaged with or without a covering liquid (Hurtado et al., 2012). Table olives, recognized as a valuable functional food for their elevated nutritional content, antioxidant properties, and abundance of beneficial components, such as dietary fiber, bioactive compounds, and monounsaturated fatty acids, hold a significant place in the Mediterranean diet (Mounir et al., 2021; Tıraş and Yıldırım, 2021). Throughout the fermentation process of these olives, LAB dominate the microbiota. This bacterial group not only enhances the organoleptic qualities of the final product but also elevates the acid content, contributing to a protective effect (Anagnostopoulos and Tsaltas, 2022; Tufariello et al., 2019; Yalçınkaya and Kılıç, 2019). The predominant genus identified in table olives is primarily Lactobacillus, but other common genera include Lactococcus, Pediococcus, Leuconostoc, and Enterococcus (Alan, 2024; Hurtado et al., 2012; Portilha-Cunha et al., 2020). Within this microbial group, enterococci and lactobacilli, in particular, are notably active in production of BAs (KučeroVá et al., 2009; Lauková et al., 2017; Zdolec et al., 2022).
Enterococcus spp. are ubiquitous microorganisms present in diverse environments, including the gastrointestinal tract (GIT) of humans and animals, as well as plants, sewage, water, soil, and various food products. These bacteria exhibit remarkable environmental persistence and resilience, demonstrating the ability to withstand a broad spectrum of temperatures and pH conditions (M’hir et al., 2012). Additionally, they can proliferate in environments containing up to 6.5% sodium chloride (NaCl) or 40% bile salts. Certain Enterococcus spp. have been utilized in the food and feed industries, functioning as starter cultures and probiotics, respectively. However, despite their beneficial applications, enterococci also contribute to food spoilage, and their presence may serve as an indicator of microbial contamination from fecal sources (Costa et al., 2022).
There is a limited research on BAs in table olives, particularly in Türkiye, and on a global scale. The distinct regional conditions exert a profound influence on the chemical composition of the final product, as their impact on LAB metabolism and their capacity for BAs production are frequently unknown (Ovalle-Marmolejo et al., 2023). Implementing control measures for BAs in traditional fermented products not only prevents food waste but also contributes to the production of healthier and higher-quality products (Akpomie et al., 2022).
This study aims to: (1) isolate Enterococcus strains from table olive samples collected in Türkiye; (2) evaluate the production capacities of these strains for tyramine, putrescine, cadaverine, histamine, and tryptamine, followed by the molecular characterization of BA-producing Enterococcus strains; and (3) quantify the levels of BAs produced by Enterococcus strains using high-performance liquid chromatography (HPLC).
Materials and Methods
Sampling
A total of 460 table olive samples—including 240 green olives and 220 black olives—were collected for analyses between August 2021 and June 2024. All samples were obtained from products fermented through spontaneous (natural) fermentation, without the use of starter cultures, in order to reflect traditional production practices and allow for the isolation of naturally occurring Enterococcus strains.
The samples were randomly collected from local markets and directly from small-scale producers across various provinces in Türkiye, representing a diverse geographical distribution. The number of samples (n) collected from each province was as follows: Mersin (65), Bursa (35), Antalya (38), Kocaeli (49), Balıkesir (47), Aydın (45), Manisa (77), Uşak (42), and Muğla (62). No commercial brand names or store-specific data were recorded, as the study aimed to capture a broad spectrum of artisanal and homemade olive products. All samples were inspected to ensure they were within their expiration dates, and were transported in portable insulated cold boxes at temperatures maintained at <4°C. Samples were not frozen at any stage. Upon arrival at the laboratory, under aseptic and refrigerated conditions, all samples were processed immediately on the same day without prior storage.
Isolation and biochemical characterization of Enterococcus spp.
To isolate enterococci, briefly, 10 g of dehyrated part of each sample was added with 90 mL of physiological saline containing 0.85% (0.1% w/v) NaCl (MerckTM, Germany) and homogenized in a stomacher (Seward 400, USA) for 8 min. This mixture was then incubated for 20 min at room temperature to ensure complete homogenization. Subsequently, serial dilutions of homogenates were prepared up to 10−5 in physiological saline, and 100 µL of each dilution was plated on Kanamycin Aesculin Azide (KAA) agar (MerckTM, Germany). Following incubation at 35–37°C for 18–24 h, three typical colonies with a black appearance on KAA were picked randomly for further identification analysis. Phenotypic characterization of all Enterococcus isolates were subjected to identification according to standard biochemical tests. These tests were Gram staining, catalase production, growth on Tryptic Soy Broth (TSB; Merck, Germany) with 6.5% NaCL, growth at pH 9.6, esculin hydrolysis on Bile Esculin Azide Agar (MerckTM, Germany) and growth at 10–45°C.
The enterococcal strains isolated in this study and reference strains were cultured on TSB and Brain Hearth Infusion (BHI) Broth (MerckTM, Germany), respectively. Incubation took place at 37°C for 24 h. The initial isolates were preserved at –20°C in 30% (v/v) aqueous glycerol (MerckTM, Germany). Three reference strains (E. faecalis ATCC 29212, Escherichia coli LMG3083 (ETEC), and Staphylococcus aureus ATCC 6538) were obtained from the culture collection of the Food Microbiology Laboratory, Department of Food Engineering, Faculty of Engineering, Ankara University, Ankara, Türkiye.
In Vitro evaluation of biogenic amine formation in Enterococcus isolates
Tyramine, putrescine, cadaverine, histamine, and tryptamine—identified as predominant BAs in olives and commonly associated with enterococcal strains—were analyzed. To evaluate the decarboxylase activity of Enterococcus spp. isolates, a modified decarboxylase medium described by Maijala (1993) was utilized. The medium was prepared with the following components per liter of distilled water: 1-g dextrose, 5-g peptone, 0.02-g bromocresol purple, and 3-g yeast extract. Amino acids corresponding to the targeted BAs (L-tyrosine, L-lysine, L-ornithine, L-tryptophan, and L-histidine) were added to the medium sequentially, each at a final concentration of 0.5%. The pH was adjusted to 6.78–6.82 using 1 N NaCl and 1 N HCl. The medium was then autoclaved at 121°C for 15 min. Separate broth tubes were prepared for each amino acid, with a control medium lacking added amino acids included for comparison.
Freshly activated bacterial cultures were inoculated into 0.1 mL of decarboxylase broth at an optical density of 0.50 at 600 nm (OD600). The cultures were then incubated at 30°C for 4–5 days, with daily monitoring to detect any color changes. In the control tube, which lacked amino acids, the medium was expected to remain yellow, indicating a negative result. A color shift from yellow to purple in the medium containing amino acid was interpreted as a positive result for BA formation, based on the criteria outlined by Bover-Cid and Holzapfel (1999).
Genotypic characterization of Enterococcus spp. isolates
Biogenic amines producing Enterococcus spp. isolates were identified by amplifying and sequencing the 16S rDNA gene. Genomic DNA was initially extracted from overnight TSB cultures of enterococcal and control strains using the GeneAll genomic DNA purification kit (Catalog No.: 106-101). DNA concentration and purity were measured spectrophotometrically with a NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific, IL, USA), and the extracted DNA was stored at –20°C. The amplification of the 16S rDNA gene utilized universal primers 907R (CCGTCAATTCMTTTRAGTTT) and 27F (AGAGTTTGATCMTGGCTCAG), as recommended by Beasley and Saris (2004).
Each 50-μL polymerase chain reaction (PCR) mixture contained 3 μL of bacterial DNA template, 34.75-μL RNase/DNase-free water, 0.25-μL Taq DNA polymerase in reaction buffer, 1 μL of 2-mM each dNTP, 4 μL of 25-mM MgCl2, 1 μL of each primer (forward and reverse), and 5 μL of PCR buffer. PCR amplifications were carried out using a Thermo Cycler (Techne TC-512, Staffordshire, UK) under the following conditions: initial denaturation at 95°C for 4 min, followed by 30 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 6 min, with a final extension step at 72°C for 8 min. The resulting PCR products were purified using the GeneJET PCR purification kit (Thermo Fisher Scientific) and analyzed via 1% agarose gel electrophoresis. The gels were stained with ethidium bromide, visualized under Ultraviolet (UV) light, and compared against an O’GeneRuler™ 10,000-bp DNA ladder (Thermo Fisher Scientific) to determine fragment sizes. The sequences obtained were analyzed using the BLAST program to compare them with the 16S rDNA sequences in the National Center for Biotechnology Information (NCBI) database.
Quantification of biogenic amine production by HPLC
Only the Enterococcus strains that tested positive for decarboxylase activity in the preliminary in vitro screening were subjected to quantitative analysis using HPLC. The quantification of BAs in TSB culture supernatants was conducted using HPLC by following the acid extraction and derivatization protocol described by Sang et al. (2020). Initially, the enterococcal bacterial strains were incubated in TSB at 37°C for 24 h. The cultures were then transferred to TSB supplemented with 0.25% histidine, lysine, tyrosine, and ornithine hydrochloride and incubated at 37°C for an additional 48 h. For sample preparation, 1 mL of the culture was mixed with 1 mL of 5% trichloroacetic acid (TCA) and centrifuged at 4°C for 10 min. Derivatization of the supernatant involved the addition of 50 μL of 2 mol/L sodium hydroxide, followed by 300 μL of 10 mg/mL dansyl chloride and 100 μL of saturated sodium bicarbonate. Subsequently, 50 μL of 25% ammonia was added, and the mixture was kept in the dark at 25°C for 30 min. The concentrations of histamine, cadaverine, tryptamine, tyramine, and putrescine were measured using a Shimadzu LC-2030 HPLC system (Kyoto, Japan) equipped with a C18 column (Agilent ZORBAX Eclipse XDB-C18, 4.6 × 250 mm, 5 μm). Mobile phases consisted of ultrapure water (phase A) and acetonitrile (phase B). The flow rate was maintained at 1 mL/min, with PDA detection at 254 nm. The analysis was conducted with a gradient elution program as follows: 0–5 min, 65–70% B; 5–14 min, 70–100% B; 14–18 min, 100% B; 18–20 min, 100–65% B; 20–22 min, 65% B. Regression parameters of BA compounds determined by the HPLC method are presented in Table 1, indicating good linearity and method reliability.
Table 1. Regression parameters of biogenic amine (BA) compounds determined by HPLC method.
| Wavelength (nm) | Regression equationy = m (x) + n | Correlation coefficient (r) | Linear rangea(mg L–1) | LODb(mg L−1) | LOQb(mg L−1) | |
|---|---|---|---|---|---|---|
| Tryptamine | 254 | y = 3.27 109 (x) – 2.15 106 | 0.9995 | 0.10–50 | 0.014 | 0.043 |
| Putrescine | 254 | y = 4.98 109 (x) + 1.08 106 | 0.9998 | 0.10–200 | 0.023 | 0.071 |
| Cadaverine | 254 | y = 3.15 107 (x) + 3.46 106 | 0.9994 | 0.10–50 | 0.008 | 0.023 |
| Histamine | 254 | y = 4.12 109 (x) + 1.72 106 | 0.9989 | 0.05–50 | 0.001 | 0.003 |
| Tyramine | 254 | y = 5.76 109 (x) – 9.85 106 | 0.9995 | 0.10–250 | 0.005 | 0.014 |
a10 calibration points were studied for linearity range (n = 10); bthree replicates were performed (n = 3).
LOD (µg/mL) = 3.3 (SD of the response/slope); LOQ (µg/mL) = 10 (SD of the response/slope).
Statistical and chemometric analysis
Statistical and chemometric analyses (principal component analysis [PCA], hierarchical cluster analysis [HCA], and Pearson’s correlation analysis) were performed using the Minitab software (version 17 for PC; Minitab Inc., UK). These were applied to data on different BA compounds. Results of statistical analysis were obtained by using multiple analyses of variance.
Nucleotide sequence accession numbers
The nucleotide sequences of the 16S rDNA genes from 71 Enterococcus isolates in this study were submitted to GenBank. The corresponding accession numbers are provided in Table 2.
Table 2. The accession numbers of Enterococcus strains used in this study.
| Strains | Accession number | Strains | Accession number |
|---|---|---|---|
| Enterococcus faecalis 2 | PV057399 | Enterococcus lactis 89 | PV091859 |
| Enterococcus faecalis 4 | PV057389 | Enterococcus lactis 103 | PV091853 |
| Enterococcus faecalis 8 | PV057397 | Enterococcus lactis 105 | PV091846 |
| Enterococcus faecalis 13 | PV057415 | Enterococcus. lactis 127 | PV091865 |
| Enterococcus faecalis 19 | PV057401 | Enterococcus lactis 139 | PV091849 |
| Enterococcus faecalis 22 | PV057393 | Enterococcus lactis 142 | PV091863 |
| Enterococcus faecalis 31 | PV057407 | Enterococcus lactis 148 | PV091856 |
| Enterococcus faecalis 36 | PV057409 | Enterococcus lactis 152 | PV091858 |
| Enterococcus faecalis 41 | PV057413 | Enterococcus lactis 163 | PV091864 |
| Enterococcus faecalis 44 | PV057391 | Enterococcus lactis 167 | PV091847 |
| Enterococcus faecalis 48 | PV057395 | Enterococcus lactis 187 | PV091851 |
| Enterococcus faecalis 50 | PV057403 | Enterococcuslactis 205 | PV091860 |
| Enterococcus faecalis 56 | PV057414 | Enterococcus lactis 214 | PV091861 |
| Enterococcus faecalis 63 | PV057390 | Enterococcus lactis 223 | PV091854 |
| Enterococcus faecalis 66 | PV057392 | Enterococcus lactis 245 | PV091850 |
| Enterococcus faecalis 88 | PV057416 | Enterococcus faecium 10 | PV056140 |
| Enterococcus faecalis 93 | PV057394 | Enterococcus faecium 40 | PV056138 |
| Enterococcus faecalis 110 | PV057396 | Enterococcus faecium 52 | PV056141 |
| Enterococcus faecalis 116 | PV057411 | Enterococcus faecium 61 | PV056139 |
| Enterococcus faecalis 122 | PV057398 | Enterococcus faecium 92 | PV056153 |
| Enterococcus faecalis 129 | PV057417 | Enterococcus faecium 98 | PV056154 |
| Enterococcus faecalis 138 | PV057400 | Enterococcus faecium 104 | PV056145 |
| Enterococcus faecalis 161 | PV057402 | Enterococcus faecium 128 | PV056156 |
| Enterococcus faecalis 185 | PV057404 | Enterococcus faecium 149 | PV056142 |
| Enterococcus faecalis 200 | PV057405 | Enterococcus faecium 158 | PV056149 |
| Enterococcus faecalis 206 | PV057406 | Enterococcus faecium 170 | PV056152 |
| Enterococcus faecalis 215 | PV057408 | Enterococcus faecium 174 | PV056155 |
| Enterococcus faecalis 227 | PV057410 | Enterococcus faecium 179 | PV056150 |
| Enterococcus faecalis 233 | PV057418 | Enterococcus faecium 193 | PV056157 |
| Enterococcus faecalis 244 | PV057419 | Enterococcus faecium 197 | PV056151 |
| Enterococcus faecalis 251 | PV057412 | Enterococcus faecium 213 | PV056144 |
| Enterococcus lactis 21 | PV091857 | Enterococcus faecium 218 | PV056147 |
| Enterococcus lactis 27 | PV091862 | Enterococcus faecium 230 | PV056148 |
| Enterococcus lactis 59 | PV091848 | Enterococcus faecium 248 | PV056146 |
| Enterococcus lactis 70 | PV091855 | Enterococcus faecium 253 | PV056143 |
| Enterococcus lactis 76 | PV091852 |
Results
Of the total of 460 spontaneously fermented table olive samples collected, 186 Enterococcus isolates were recovered, corresponding to an overall isolation rate of 40.43% (data not shown). Morphological and cultural tests were applied to 186 enterococcal isolates. All of the isolates showed developmental characteristics at pH 9.6, 6.5% NaCL, and at 10–45°C. In addition, these isolates were also identified as Gram-positive, catalase negative, and esculin hydrolysis positive. Among the 186 Enterococcus strains, 71 (38.17%) were identified as BA producers. In all, 71 isolates isolated from 50 green and 21 black olives were identified at species level by 16S rDNA sequence analysis (Figure 1). Enterococcus strains were identified as: 20 E. faecium (28.17%), 31 E. faecalis (43.66%), and 20 E. lactis (28.17%). The strains of E. faecium and E. lactis were isolated from 15 green and 5 black olives, while the strains of E. faecalis were isolated from 20 green and 11 black olives (Tables 3 and 4).
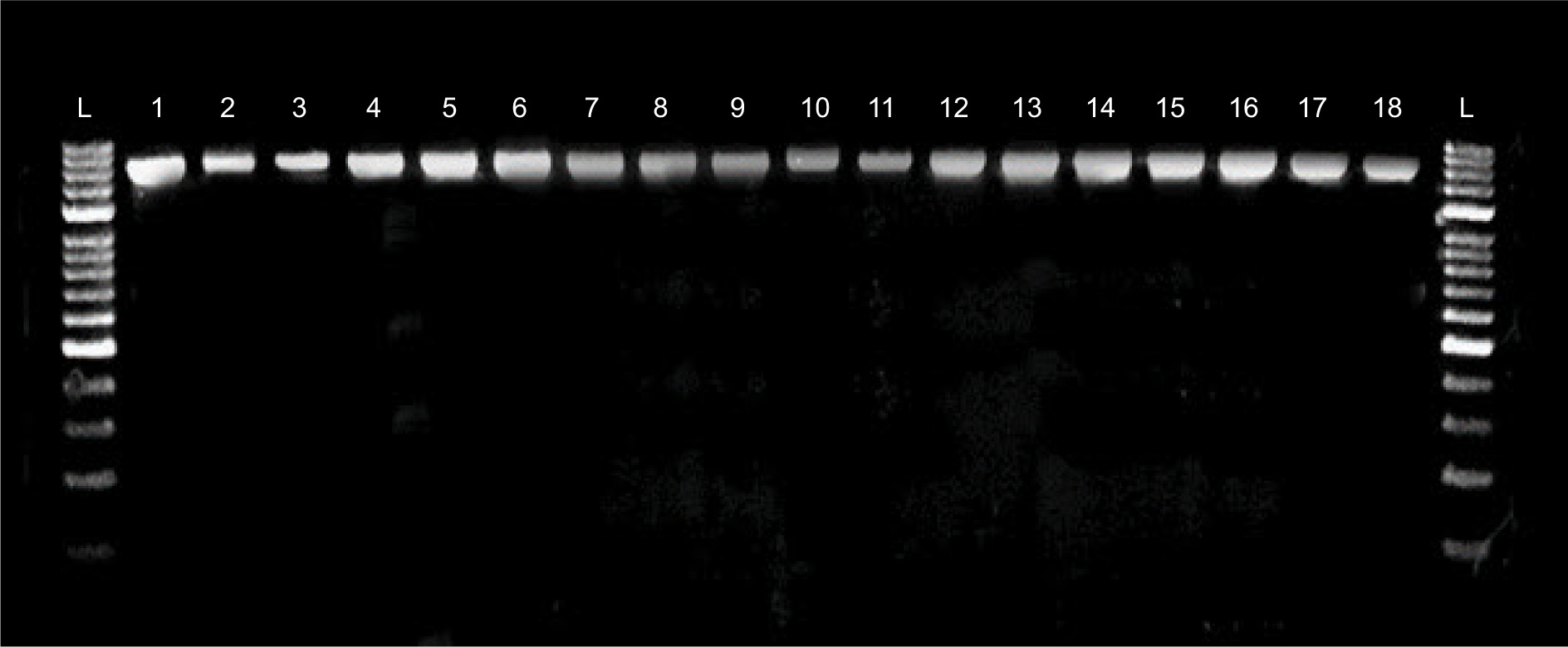
Figure 1. 16S rDNA fragments of Enterococcus spp. strains. L: O’Gene ruler DNA marker. 1. E. lactis 139; 2. E. faecium 40; 3. E. faecium 10; 4. E. faecalis 4; 5. E. faecalis 44; 6. E. faecalis 22; 7. E. faecalis 48; 8. E. faecalis 8; 9. E. lactis 27; 10. E. faecalis 2; 11. E. faecalis 19; 12. E. faecalis 50; 13. E. faecium 52; 14. E. faecalis 200; 15. E. faecalis 31; 16. E. faecalis 36; 17. E. lactis 76; 18. E. faecalis 116.
Table 3. Concentrations of BAs (mg/L) produced by Enterococcus spp. strains isolated from green fermented olives.
| Strains | Tyramine | Cadaverine | Putrescine | Tyrptamine | Histamine | Total BAs |
|---|---|---|---|---|---|---|
| E. faecalis 2 | 191.346±0.987 | 0.593±0.009 | 0.400±0.030 | ND | ND | 192.339±0.988 |
| E. faecalis 4 | 184.531±1.021 | 0.549±0.011 | ND | 0.640±0.042 | 1.158±0.027 | 186.878±1.022 |
| E. faecalis 8 | 1.363±0.005 | ND | 0.426±0.031 | 1.355±0.011 | 2.356±0.046 | 5.500±0.057 |
| E. faecalis 41 | 32.057±0.411 | 1.280±0.044 | 139.620±0.865 | 3.684±0.067 | 4.645±0.032 | 181.286±0.962 |
| E. faecalis 44 | 15.789±0.023 | 2.026±0.032 | 46.997±0.499 | ND | ND | 64.812±0,501 |
| E.faecalis 48 | 52.185±0.564 | 3.564±0.065 | 21.604±0.234 | 2.284±0.040 | 0,873±0.009 | 80.510±0.615 |
| E. faecalis 50 | 16.004±0.042 | 2.340±0.054 | 23.811±0.211 | 0.462±0.028 | 5.647±0.071 | 48.264±0.235 |
| E. faecalis 63 | 1.279±0.023 | ND | ND | 1.466±0.032 | 7.985±0.096 | 10.730±0.104 |
| E. faecalis 66 | 30.739±0.387 | ND | ND | ND | 4.196±0.065 | 34.935±0.392 |
| E. faecalis 93 | 40.822±0.396 | 0.831±0.008 | 54.285±0.632 | ND | 2.015±0.045 | 97.953±0.747 |
| E. faecalis 110 | ND | ND | ND | 1.546±0.086 | ND | 1.546±0.086 |
| E. faecalis 122 | 0.691±0.004 | ND | ND | 1.133±0.054 | ND | 1.824±0.054 |
| E. faecalis 138 | 0.756±0.005 | ND | ND | 0.871±0.023 | ND | 1.627±0.024 |
| E. faecalis 161 | 21.917±0.119 | ND | ND | 1.601±0.043 | ND | 23.518±0.127 |
| E. faecalis 185 | 15.113±0.032 | 0.886±0.008 | 48.781±0.498 | ND | 0.156±0.009 | 64.936±0.499 |
| E. faecalis 200 | 58.158±0.514 | ND | 30.314±0.315 | 2.558±0.111 | 0.546±0.007 | 91.576±0.613 |
| E. faecalis 206 | 202.254±1.102 | ND | ND | 0.761±0.009 | 1.218±0.044 | 204.233±1.103 |
| E. faecalis 215 | 257.939±1.654 | ND | ND | 1.312±0.042 | ND | 259.251±1.656 |
| E. faecalis 227 | 35.242±0.298 | 13.923±0.067 | 3.094±0.025 | 4.058±0.067 | ND | 56.317±0.314 |
| E. faecalis 251 | 16.367±0.054 | 3.904±0.013 | 35.853±0.398 | 0.536±0.010 | ND | 56.660±0.402 |
| E. lactis 21 | 1.127±0.034 | ND | ND | 1.420±0.050 | ND | 2.547±0.061 |
| E. lactis 27 | 202.099±1.244 | ND | ND | 2.974±0.065 | 0.547±0.007 | 205.620±1.246 |
| E. lactis 70 | 21.097±0.124 | ND | ND | 2.246±0.059 | 1.687±0.031 | 25.030±0.141 |
| E. lactis 89 | 21.953±0.132 | ND | ND | 0.717±0.010 | 0.978±0.010 | 23.648±0.133 |
| E. lactis 103 | 15.655±0.098 | ND | ND | 0.638±0.009 | ND | 16.293±0.098 |
| E. lactis 127 | 2.800±0.035 | 1.595±0.037 | 0.485±0.029 | ND | ND | 4.880±0.059 |
| E. lactis 142 | 3.765±0.056 | 1.570±0.033 | 0.671±0.017 | ND | ND | 6.006±0.067 |
| E. lactis 148 | ND | ND | 11.562±0.087 | 25.183±0.125 | ND | 36.745±0.152 |
| E. lactis 152 | 3.056±0.063 | 1.696±0.043 | 0.598±0.031 | ND | ND | 5.350±0.082 |
| E. lactis 163 | ND | ND | ND | 10.260±0.044 | 6.285±0.072 | 16.545±0.084 |
| E. lactis 167 | 1.029±0.005 | ND | ND | 1.501±0.032 | ND | 2.530±0.032 |
| E. lactis 187 | 0.678±0.023 | ND | 1.387±0.101 | 2.826±0.057 | ND | 4.891±0.118 |
| E. lactis 205 | ND | ND | 12.814±0.097 | 16.120±0.101 | 0.126±0.009 | 29.060±0.140 |
| E. lactis 214 | ND | ND | ND | ND | 1.018±0.044 | 1.018±0.044 |
| E. lactis 223 | 226.676±2.001 | 7.997±0.654 | ND | 24.651±0.265 | ND | 259.324±2.122 |
| E. faecium 40 | 21.386±0.234 | 4.295±0.023 | 49.304±0.401 | 30.562±0.301 | ND | 105.547±0.554 |
| E. faecium 52 | 52.797±0.899 | 2.936±0.016 | 13.518±0.088 | 1.851±0.034 | ND | 71.102±0.899 |
| E. faecium 61 | 1.001±0.008 | ND | ND | 1.452±0.027 | 1.569±0.054 | 4.022±0.061 |
| E. faecium 92 | 0.448±0.034 | ND | ND | 0.995±0.019 | ND | 1.443±0.039 |
| E. faecium 98 | 22.965±0.119 | ND | 93.817±0.897 | ND | ND | 116.782±0.905 |
| E. faecium 128 | 0.988±0.007 | ND | ND | 1.504±0.031 | ND | 2.492±0.008 |
| E. faecium 149 | 14.817±0.119 | ND | ND | 2.128±0.047 | ND | 16.945±0.128 |
| E. faecium 158 | 1.149±0.008 | ND | ND | 1.625±0.039 | ND | 2.774±0.040 |
| E. faecium 170 | 72.093±0.883 | 0.990±0.008 | ND | 1.102±0.029 | ND | 74.185±0.884 |
| E. faecium 174 | 207.265±2.105 | ND | ND | 2.608±0.049 | ND | 209.873±2.106 |
| E. faecium 179 | 0.876±0.035 | ND | ND | 1.796±0.037 | ND | 2.672±0.051 |
| E. faecium 193 | 0.873±0.041 | ND | ND | 2.075±0.041 | ND | 2.948±0.058 |
| E. faecium 197 | 0.865±0.043 | ND | ND | 3.647±0.054 | ND | 4.512±0.069 |
| E. faecium 248 | 13.933±0.098 | 4.018±0.032 | 51.343±0.675 | ND | ND | 69.294±0.682 |
| E. faecium 253 | 4.932±0.054 | 1.197±0.010 | 0.469±0.031 | 0.798±0.011 | 0.159±0.001 | 7.555±0.064 |
ND: not determined; BAs: biogenic amines;
(n = 3, all parameters are given with their standard deviations).
Table 4. Concentrations of biogenic amines (BAs; mg/L) produced by Enterococcus spp. strains isolated from black fermented olives.
| Strains | Tyramine | Cadaverine | Putrescine | Tyriptamine | Histamine | Total BAs |
|---|---|---|---|---|---|---|
| E. faecalis 13 | 207.618±1.021 | ND | ND | 2.572±0.027 | ND | 210.190±1.021 |
| E. faecalis 19 | 136.999±0.980 | 3.528±0.053 | 55.355±0.547 | 1.019±0.013 | ND | 196.901±1.124 |
| E. faecalis 22 | 17.960±0.025 | 2.732±0.044 | 25.726±0.123 | ND | ND | 46.418±0.133 |
| E. faecalis 31 | 88.080±0.654 | 3.849±0.031 | 121.696±0.879 | 1.053±0.003 | ND | 214.678±1.096 |
| E. faecalis 36 | 29.245±0.167 | 1.403±0.024 | 78.437±0.654 | 1.152±0.004 | ND | 110.237±0.675 |
| E. faecalis 56 | 16.121±0.031 | 2.230±0.022 | 36.702±0.345 | ND | ND | 55.053±0.347 |
| E. faecalis 88 | 1.172±0.004 | ND | ND | 1.514±0.007 | ND | 2.686±0.008 |
| E. faecalis 116 | 34.567±0.245 | ND | 148.718±1.214 | 9.095±0.023 | ND | 192.380±1.239 |
| E. faecalis 129 | ND | ND | ND | 22.785±0.121 | ND | 22.785±0.121 |
| E. faecalis 233 | 1.145±0.024 | ND | ND | 1.539±0.009 | ND | 2.684±0.026 |
| E. faecalis 244 | 0.468±0.006 | ND | ND | 1.363±0.009 | ND | 1.831±0.011 |
| E. lactis 59 | 5.412±0.065 | 1.286±0.075 | 0.617±0.021 | ND | ND | 7.315±0.102 |
| E. lactis 76 | 37.170±0.315 | ND | 122.530±1.021 | 0.548±0.004 | ND | 160.248±1.069 |
| E. lactis 105 | 1.090±0.098 | ND | ND | 0.751±0.003 | ND | 1.841±0.098 |
| E. lactis 139 | 35.078±0.411 | 0.989±0.043 | 53.708±0.542 | ND | ND | 89.775±0.682 |
| E. lactis 245 | 13.178±0.019 | ND | ND | 0.870±0.007 | ND | 14.048±0.020 |
| E. faecium 10 | 107.251±0.998 | 0.815±0.032 | ND | 1.160±0.008 | ND | 109.226±0.999 |
| E. faecium 104 | 3.888±0.012 | 1.436±0.081 | 0.637±0.019 | 0.374±0.000 | ND | 6.335±0.084 |
| E. faecium 213 | 3.312±0.021 | 1.852±0.078 | 0.568±0.023 | ND | ND | 5.732±0.084 |
| E. faecium 218 | 21.208±0.301 | ND | ND | 1.943±0.098 | ND | 23.151±0.317 |
| E. faecium 230 | 1.057±0.087 | ND | ND | 1.477±0.011 | ND | 2.534±0.088 |
ND: not determined; BAs: biogenic amines;
(n = 3, all parameters are given with their standard deviations).
Enterococcus strains isolated from fermented green olives (G) were identified as producers of tyramine (45 strains; TyrG), tryptamine (39 strains; TypG), putrescine (22 strains; PutG), cadaverine (19 strains; CadG), and histamine (19 strains; HisG). The concentrations of TyrG, CadG, PutG, TypG, and HisG in the samples were determined to range from ND to 257.939±1.654 mg/L, ND to 13.923±0.067 mg/L, ND to 139.620±0.865 mg/L, ND to 30.562±0.301 mg/L, and ND to 7.985±0.096 mg/L, respectively (Table 3). The total BA (TotG) content produced by the Enterococcus strains isolated from green olives was in the range of 1.018±0.044–259.324±2.122 mg/L. The TotG content produced by E. faecalis strains ranged from 1.546±0.086 mg/L to 259.251±1.656 mg/L, while that produced by E. lactis strains ranged from 1.018±0.044 mg/L to 259.324±2.122 mg/L, and E. faecium strains produced amounts ranging from 1.443±0.039 mg/L to 209.873±2.106 mg/L. The concentrations of TyrG, CadG, PutG, TypG, and HisG produced by E. faecalis strains were found in the range of ND–257.939±1.654 mg/L, ND–13.923±0.067 mg/L, ND–139.620±0.865 mg/L, ND–4.058±0.067 mg/L, and ND–7.985±0.096 mg/L, respectively. For E. lactis strains, the concentrations of TyrG, CadG, PutG, TypG, and HisG ranged from ND to 226.676±2.001 mg/L, ND to 7.997±0.654 mg/L, ND to 12.814±0.097 mg/L, ND to 25.183±0.125 mg/L, and ND to 6.285±0.072 mg/L, respectively. Similarly, E. faecium strains produced TyrG, CadG, PutG, TypG, and HisG at concentrations ranging from 0.448 to 207.265±2.105 mg/L, ND to 4.295±0.023 mg/L, ND to 93.817±0.897 mg/L, ND to 30.562±0.301 mg/L, and ND–1.569±0.054 mg/L, respectively.
In all, 20 Enterococcus strains isolated from fermented black olives (B) were identified as tyramine (TyrB) producers, 10 as cadaverine (CadB) producers, 11 as putrescine (PutB) producers, and 16 as tryptamine (TypB) producers. However, none of the Enterococcus strains were found to produce histamine (HisB). The concentrations of TyrB, CadB, PutB, and TypB in the samples were determined to range from ND to 207.618±1.021 mg/L, ND to 3.849±0.031 mg/L, ND to 148.718±1.214 mg/L, and ND to 22.785±0.121 mg/L, respectively (Table 4). The total BA (TotB) content generated by the Enterococcus strains isolated from black olives ranged from 1.831±0.011 mg/L to 214.678±1.096 mg/L. TotB production by E. faecalis strains was measured as 1.831±0.011–214.678±1.096 mg/L, while E. lactis strains produced 1.841±0.098–160.248±1.069 mg/L, and E. faecium strains produced 2.534±0.088–109.226±0.999 mg/L. TyrB, CadB, PutB, and TypB levels produced by E. faecalis strains were reported within the range of ND–207.618±1.021 mg/L, ND–3.849±0.031 mg/L, ND–148.718±1.214 mg/L, and ND–22.785±0.121 mg/L, respectively. In E. lactis strains, the respective concentrations of TyrB, CadB, PutB, and TypB ranged from 1.090±0.098 mg/L to 37.170±0.315 mg/L, ND to 1.286±0.075 mg/L, ND to 122.530±1.021 mg/L, and ND to 0.870±0.007 mg/L. Similarly, E. faecium strains produced TyrB, CadB, PutB, and TypB in the range of 1.057±0.087–107.251±0.998 mg/L, ND–1.852±0.078 mg/L, ND–0.6370.019 mg/L, and ND–1.943±0.098 mg/L, respectively.
The most abundantly produced BAs in green olives were identified as TyrG (257.939±1.654 mg/L, produced by E. faecalis 215), PutG (139.620±0.865 mg/L, produced by E. faecalis 41), TypG (30.562±0.301 mg/L, produced by E. faecium 40), CadG (13.923±0.067 mg/L, produced by E. faecalis 227), and HisG (7.985±0.096 mg/L, produced by E. faecalis 63). In black olives, the highest levels of BAs were recorded as TyrB (207.618±1.021 mg/L, produced by E. faecalis 13), PutB (148.718±1.214 mg/L, produced by E. faecalis 116), TypB (22.785±0.121 mg/L, produced by E. faecalis 129), and CadB (3.849±0.031 mg/L, produced by E. faecalis 31).
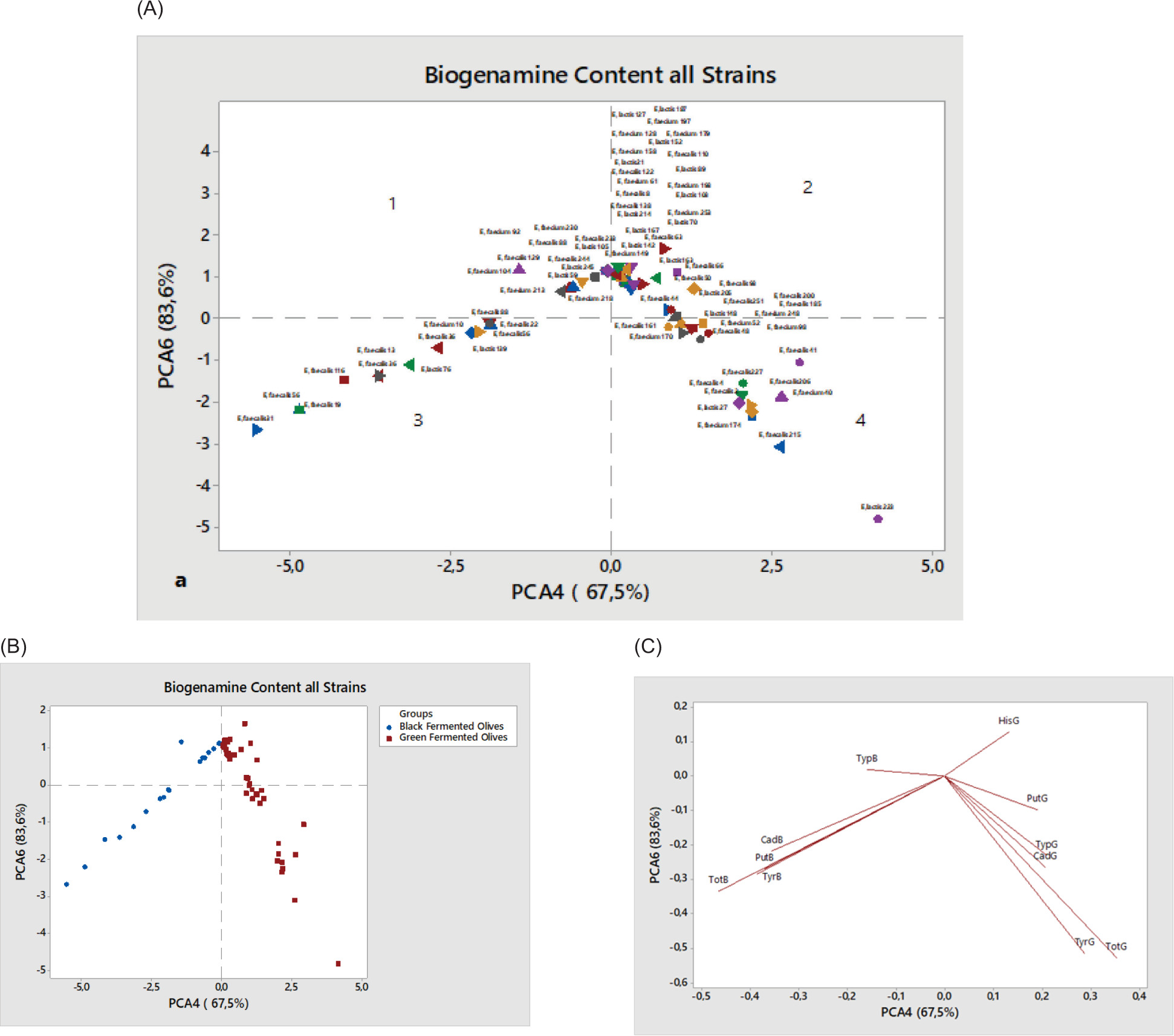
In order to gain a more comprehensive understanding of the trends and relationships among the examined variables in relation to BA composition in traditionally fermented green and black table olives, PCA was performed (Cheng et al., 2010). The first four principal components (PCs) accounted for more than 83.60% of the total variance, with the first two PCs explaining approximately 57.60% of the observed variability. The fourth PC (PC4), which represented 67.50% of the total variance, was positively associated with HisG, showing negative correlations with PutG, TypG, CadG, TyrG, and TotG (Figure 2C, regions 2 and 4). The sixth PC (PC6) accounted for 16.10% of the total variance and was positively correlated with TypB whereas it was negatively associated with CadB, PutB, TyrB, and TotB (Figure 2C, regions 1 and 3).
Figure 2. (A) Principal component analysis (PCA) score plot, and (B and C) loading plots for biogenic amines in table olive oils.
The PCA results indicated that PC4 and PC6 effectively differentiated two distinct groups of table olives. The first group consisting of black fermented olives was positioned on the left side (regions 1 and 3), while the second group, composed of green fermented olives, was located on the right side (regions 2 and 4) (Figure 2B). A combined analysis of Figures 2A and 2C revealed that 12 strains in region 1—E. faecium 92, E. faecium 230, E. faecalis 88, E. faecalis 233, E. lactis 105, E. faecalis 244, E. faecium 104, E. faecalis 129, E. lactis 245 , E. lactis 59, E. faecium 213, and E. faecium 218—were positively associated with TypB. Similarly, in region 3, strains E. faecalis 31, E. faecalis 56, E. faecalis 19, E. faecalis 116 , E. faecalis 13, E. faecalis 36, E. lactis 76, E. lactis 139, E. faecalis 22, E. faecalis 88, and E. faecium 10 exhibited negative associations with CadB, PutB, TyrB, and TotB. In region 4, strains E. faecalis 161, E. faecium 170, E. faecium 52, E. faecium 98, E. faecalis 48, E. faecalis 41, E. faecalis 227, E. faecalis 4, E. faecalis 2, E. lactis 27, E. faecium 174, E. faecalis 215, E. faecalis 206, E. faecium 40, and E. lactis 223 displayed negative correlations with PutG, TypG, CadG, TyrG, and TotG whereas strains in region 2 exhibited positive associations with HisG.
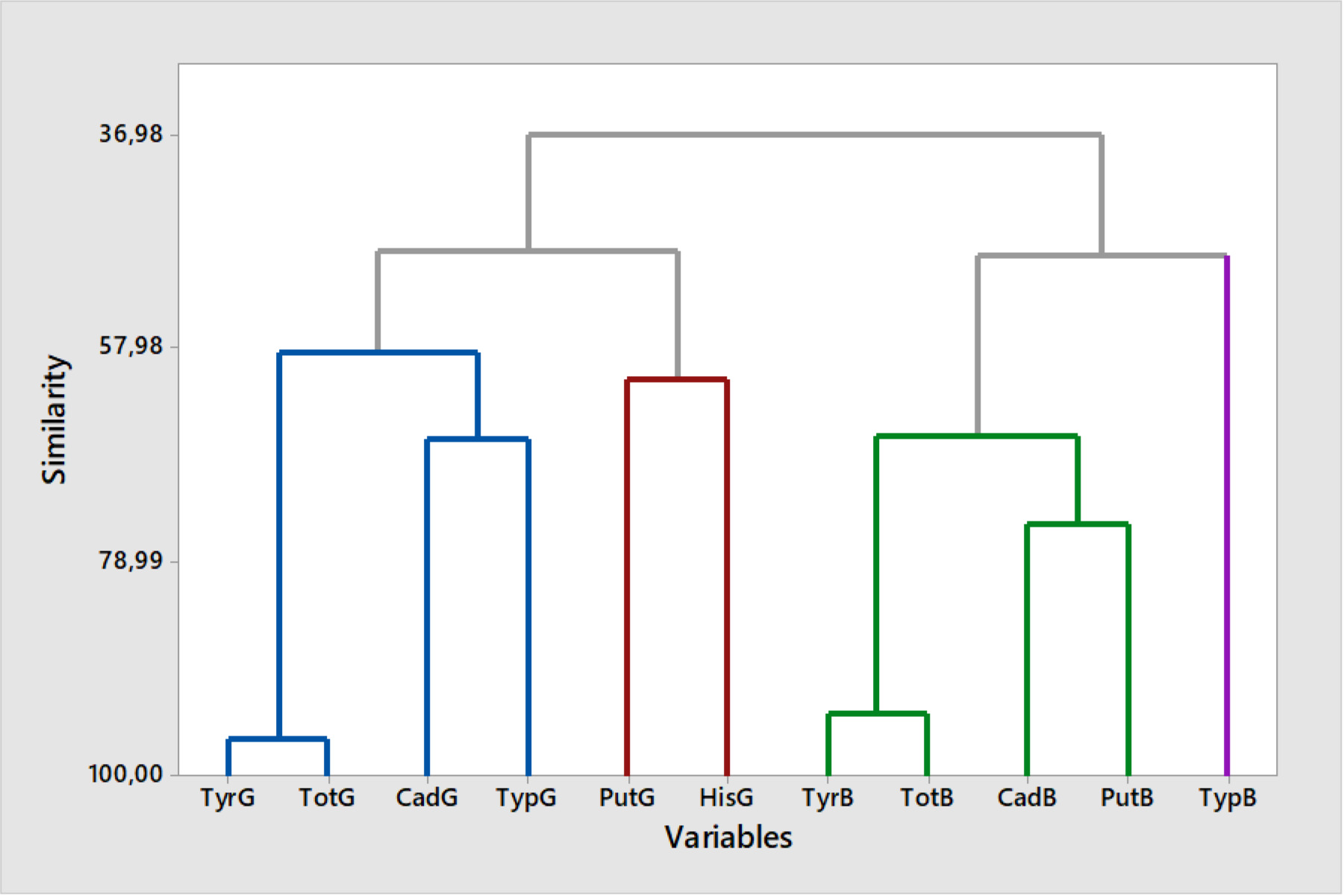
A clustering analysis was performed using the non-hierarchical k-means method, resulting in 11 components (Figure 3). This approach grouped each variable into clusters based on their similarity, with the clustering process visualized in a dendrogram. The analysis revealed that 11 variables were categorized into four main interrelated clusters: (i) cluster 1 included four components (TyrG, CadG, TypG, and TotG), (ii) cluster 2 consisted of two components (PutG and HisG), which were closely related, indicating similar characteristics, (iii) cluster 3 encompassed four components (TyrB, CadB, PutB, and TotB), and (iv) cluster 4 contained a single component (TrpB). The strongest correlation (96.31%) was observed between TyrG and TotG, while CadG and TypG exhibited a lower similarity (66.78%). The overall relationship among the four components in cluster 1 was determined to be 58.35%. Among the least similar variables, TypB exhibited a similarity of 48.82%. Within cluster 3, the closest association was observed between TyrB and TotB (94.02% similarity) whereas CadB and PutB demonstrated a relatedness of 75.29%. The overall connectivity of the binary groups in cluster 3 was 66.62%.
Figure 3. Dendrogram of biogenic amines in table olive oils using the non-hierarchical k-means method based on squared Euclidean distance across all strains.
Pearson’s correlation analysis was conducted to assess the relationships among BA compounds. Statistically significant correlations were identified between TotG and TyrG (r = 0.926, p < 0.000), TotB and TyrB (r = 0.880, p < 0.000), TotB and PutB (r = 0.738, p < 0.000), and TotB and CadB (r = 0.563, p < 0.000). No other statistically significant correlations were observed among the remaining variables.
Discussion
This is the first comprehensive report on the characterization of Enterococcus spp. from fermented table olives in Türkiye. However, limited global information is available on the isolation of enteroccocci from table olive samples. The results obtained in this study are significant as they contribute to similar future studies. In this study, 460 table olive samples were analyzed for the presence of enterococci, and the isolation rate was determined as 40.43%. Of the 186 enterococcal strains, 38.17% were identified as BA producers.
Different isolation proportions of Enterococcus strains from fermented table olive samples were reported in the previous studies. These studies reported that the proportion of positive samples of enterococci in table olive samples was 32.22% in Western Algeria (Mourad and Nour-Eddine, 2006), 40% in Tunusia (Rehaiem et al., 2016), 69.56% in Cyprus (Anagnostopoulos et al., 2018), and 84.21% in Morocco (El Issaoui et al., 2022). However, to the best of our knowledge, only one study in Türkiye focused on the isolation of enterococci from fermented table olive samples. In that study, Yalçınkaya and Kılıç (2019) analyzed table olive samples collected from various regions of Türkiye, including Antalya, Burdur, İzmir, Isparta, and Eskişehir, and identified 9.58% of the isolates as E. faecium.
In the current study, the most prevalent species of enterococci identified was E. faecalis (43.66%), followed by E. faecium (28.17%) and E. lactis (28.17%). However, in contrast to our results, Mourad and Nour-Eddine (2006) identified E. faecium (11.6%) as the most predominant species among enterococci isolated from table olives, followed by E. faecalis (7.8%) and E. durans (7.5%). Specifically, the E. casseliflavus group represented approximately 90% of the isolates identified (34 out of 38) (De Bellis et al., 2010). Similarly, Rehaiem et al. (2016) found E. faecium to be the most common species (46.15%), with E. faecalis (27.27%), E. casseliflavus (12.58%), E. durans (8.39%), and E. mundtii (5.59%) being in prevalence. In a more recent study conducted by Anagnostopoulos et al. (2018), all 64 isolates from table olives were identified as E. faecium (68.08%).
The microbial groups in table olive fermentation primarily consisted of LAB and yeasts. The main LAB genera found in table olives include Lactobacillus, Enterococcus, Pediococcus, Leuconostoc, and Lactococcus (Albayrak and Kamber, 2020). E. faecalis and E. faeciumare recognized as frequent contaminants in fermented table olives. In addition, species such as E. casseliflavus and E. italicus are detected as part of the native microbiota during the early stages of table olive fermentation (M’hir et al., 2012). In our study, the high isolation proportion of E. faecalis and E. faecium from table olive samples was of concern. This situation was considered an indicator of a lack of hygiene.
Biogenic amines are primarily produced via the enzymatic decarboxylation of precursor amino acids, a process facilitated by microbial activity. Various species of LAB, including Enterococcus spp., Lactobacillus spp., and Pediococcus spp., which are frequently associated with spoilage in fermented food products, such as table olives, sausage, and cheese, are identified as significant producers of BAs (Liu et al., 2013; Vinci and Maddoloni, 2020). To the best of our knowledge, this is the first report on the determination of BAs producing Enterococcus spp. Strains from table olive samples in Türkiye. In our study, tyramine (90%, 45 strains) was by far the most abundant BA, with a mean of 257.939±1.654 mg/L in fermented green table olives, followed by tryptamine (78%, 39 strains), putrescine (44%, 22 strains), cadaverine (38%, 19 strains), and histamine (38%, 19 strains).
In addition, 20 Enterococcus strains isolated from fermented black olives were identified as tyramine producers (95.24%), 16 as tryptamine producers (76.19%), 11 as putrescine producers (52.38%), and 10 as cadaverine producers (47.62%). It is interesting that none of the Enterococcus strains isolated from black table olive has the ability to produce histamine. Another significant aspect of the study is that our study represents the first global report to determine the amount of BAs produced by E. lactis strains. In green table olive samples, the highest total BA production potential was detected in E. lactis strains (259.324±2.122 mg/L).
In black table olive samples, E. faecalis strains were identified as having the highest total BA production potential, with a value of 214.678±1.096 mg/L. Tyramine is the primary BA accumulated by Enterococcus spp. in substantial quantities, followed by other amines, such as putrescine, 2-phenylethylamine, cadaverine, and histamine (Houicher et al., 2024). In our study, the tyramine production potential of Enterococcus strains isolated from both green and black olives was found to be higher, compared to other BAs. Moreover, tyramine biosynthesis is a species-specific characteristic of E. faecalis, E. faecium, and E. durans, while putrescine biosynthesis was identified as a species-level trait exclusive to E. faecalis (Sun et al., 2023). However, in our study, the majority of isolated E. faecalis, E. lactis, and E. faecium strains were identified as producers of tyramine and putrescine.
In the present study, the occurrence of multiple BAs in table olive was definitively identified, with 95.77% of Enterococcus strains found to produce two or more BAs simultaneously, while only three strains were identified as producers of a single BA. In addition, among the green olive samples, 19 strains were determined to produce 2 BAs, 16 strains produced 3 BAs, 9 strains produced 4 BAs, and 4 strains produced 5 BAs. In the black olive samples, 8 strains were identified as producers of 2 BAs, 8 strains produced 3 BAs, and 4 strains produced 4 BAs. These findings are consistent with the results reported by Vesković-Moračanin et al. (2022), Yilmaz (2024), and Zhang et al. (2022).
At both European and international levels, regulatory frameworks established maximum permissible concentration limits exclusively for histamine in fish and fish-derived products. In contrast, for other food matrices, only proposed or recommended limits exist, rather than legally binding thresholds. Furthermore, no national legislation currently stipulates specific limits for other BAs or their presence in food products (FAO/WHO, 2013). No official criteria of maximum acceptable BAs concentration limits for table olives are available. Therefore, we were unable to compare the data obtained in this study based on the established criteria. However, various researchers have proposed upper concentration limits for BAs in food, including histamine at 100 mg/kg, tyramine ranging from 100 to 800 mg/kg, and total BAs at 1,000 mg/kg (Lee et al., 2024). Moreover, according to EFSA (2011), histamine-related symptoms generally manifest at exposure levels ranging from 25 to 50 mg, while histamine poisoning is reported to occur following the ingestion of 75–300 mg. The concentrations of BA obtained in the current study were below these values.
Conclusions
This study investigates the production of BAs by Enterococcus strains isolated from table olives in Türkiye, shedding light on the potential health risks associated with their presence in fermented foods. The results highlight the ability of certain Enterococcus strains to produce significant level of BAs, emphasizing the need for careful monitoring and control during the fermentation process. The findings contribute to the understanding of microbial activities in traditional table olive fermentation and provide valuable data for the development of strategies to minimize BA production in these products. This research is particularly important for safeguarding public health and ensuring the quality and safety of table olives, which hold cultural and economic significance in Türkiye as well as globally. This study not only advances our knowledge about the role of Enterococcus strains in the production of BAs but also underscores the importance of implementing microbiological and technological interventions to improve food safety standards in fermented olive production.
Availability of Data and Materials
The nucleotide sequences of the 16S rDNA gene from 71 Enterococcus isolates analyzed in the present study were submitted to and archived in GenBank. All data were included in the manuscript.
Competing Interests
The authors declared that they had no competing interests.
Author Contributions
Gülsüm Atasoy: methodology, data curation, and writing—original draft; Pınar Şanlıbaba: conceptualization, methodology, data curation, software, writing—review and editing, and supervision; Nilüfer Vural: software and writing—review; Rahmi Ertan Anlı: conceptualization, software, writing—review & editing, supervision, and funding acquisition.
Conflicts of Interest
The author declares no conflict of interest associated with this work.
Funding
This work was supported by the Ankara University Scientific Research Projects Coordination Unit (Project No. 21L0443014).
REFERENCES
Ahangari, H., Kurbanoglu, S., Ehsani, A., and Uslu, B. 2021. Latest trends for biogenic amines detection in foods: enzymatic biosensors and nanozymes applications. Trends Food Sci Technol. 112: 75–87. 10.1016/j.tifs.2021.03.037
Akpomie, O.O., Ejechi, B.O., Banach, A.M., Adewuyi, I., Ayobola, E.D., Akpomie, K.G., and Ahmadi, S. 2022. Biogenic amine production from processed animal and plant protein-based foods contaminated with Escherichia coli and Enterococcus feacalis. J Food Sci Technol. 59(12): 4880–4888. 10.1007/s13197-022-05576-0
Alan, Y. 2024. Chemical changes of potential probiotic Lactiplantibacillus plantarum and Lactobacillus pentosus starter cultures in natural Gemlik type black olive fermentation. Food Chem. 434: 137472. 10.1016/j.foodchem.2023.137472
Albayrak, Ç.B., and Kamber, A. 2020. Microflora of naturally fermented table olives and characterization of their lactic acid bacteria. ADU Ziraat Derg. 17(1): 45–52. 10.25308/aduziraat.655257
Anagnostopoulos, D.A., Bozoudi, D., and Tsaltas, D. 2018. Enterococci isolated from cypriot green table olives as a new source of technological and probiotic properties. Fermentation 4(2): 48. 10.3390/fermentation4020048
Anagnostopoulos, D.A., and Tsaltas, D. 2022. Current status, recent advances, and main challenges on table olive fermentation: the present meets the future. Front Microbiol. 12: 797295. 10.3389/fmicb.2021.797295
Banicod, R.J.S., Ntege, W., Njiru, M.N., Abubakar, W.H., Kanthenga, H.T., Javaid, A., and Khan, F. 2025. Production and transformation of biogenic amines in different food products by the metabolic activity of the lactic acid bacteria. Int J Food Microbiol. 428: 110996. 10.1016/j.ijfoodmicro.2024.110996
Barbieri, F., Montanari, C., Gardini, F., and Tabanelli, G. 2019. Biogenic amine production by lactic acid bacteria: a review. Foods 8(1): 17. 10.3390/foods8010017
Beasley, S.S., and Saris, P.E. 2004. Nisin-producing Lactococcus lactis strains isolated from human milk. Appl Environ Microbiol. 70(8): 5051–5053. 10.1128/AEM.70.8.5051-5053.2004
Bover-Cid, S., and Holzapfel, W.H. 1999. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int J Food Microbiol. 53(1): 33–41. 10.1016/S0168-1605(99)00152-X
Cheng, X.-m., Zhao, T., Yang, T., Wang, C.-h., Bligh, S.W.A., and Wang, Z.-t. 2010. HPLC fingerprints combined with principal component analysis, hierarchical cluster analysis and linear discriminant analysis for the classification and differentiation of Peganum spp. indigenous to China. Phytochem Anal. 21: 279–289. 10.1002/pca.1198
Costa, L.D.F., Falcao, D.A., Grassotti, T.T., Christiano, F.D., Frazzon, J., and Frazzon, A.P.G. 2022. Antimicrobial resistance of enterococci isolated from food in South Brazil: Comparing pre-and post-RDC 20/2011. Ann Acad Bras de Ciênc. 94(1): e20201765. 10.1590/0001-3765202220201765
De Bellis, P., Valerio, F., Sisto, A., Lonigro, S.L., and Lavermicocca, P. 2010. Probiotic table olives: microbial populations adhering on olive surface in fermentation sets inoculated with the probiotic strain Lactobacillus paracasei IMPC21 in an industrial plant. Int J Food Microbiol. 140(1): 6–13. 10.1016/j.ijfoodmicro.2010.02.024
El Issaoui, K., Senhaji, N.S., Wieme, A., Abrini, J., and Khay, E.O. 2022. Probiotic properties and physicochemical potential of lactic acid bacteria ısolated from Moroccan table olives. J Food Qual Hazards Control. 9(3): 169–178. 10.18502/jfqhc.9.3.11155
European Food Safety Authority (EFSA). 2011. Scientific opinion on risk-based control of biogenic amine formation in fermented foods. Panel on Biological Hazards (BIOHAZ). Efsa J. 9(10): 2393–2487. 10.2903/j.efsa.2011.2393
Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO). 2013. Joint FAO/WHO Expert Meeting on the Public Health Risks of Histamine and Other Biogenic Amines from Fish and Fishery Products: Meeting Report, 23–27 July 2012. FAO/WHO, Rome, Italy. ISBN 978-92-5-107849-5
Gao, P., Mohd Noor, N.Q.I., and Md. Shaarani, S. 2022. Current status of food safety hazards and health risks connected with aquatic food products from Southeast Asian region. Crit Rev Food Sci Nutr. 62(13): 3471–3489. 10.1080/10408398.2020.1866490
Ghorbani, M., Molaei, R., Moradi, M., Tajik, H., Salimi, F., Kousheh, S.A., and Koutamehr, M.E. 2021. Carbon dots-assisted degradation of some common biogenic amines: an in vitro study. Food Sci Technol (LWT). 136: 110320. 10.1016/j.lwt.2020.110320
Guba, A., Bába, O., Tőzsér, J., Csősz, É., and Kalló, G. 2022. Fast and sensitive quantification of AccQ-tag derivatized amino acids and biogenic amines by UHPLC-UV analysis from complex biological samples. Metabolites 12(3): 272. 10.3390/metabo12030272
Houicher, A., Kuley, E., Bensid, A., Yazgan, H., and Özogul, F. 2024. In vitro study of biogenic amine production and gastrointestinal stress tolerance by some enterococci strains. J Sci Food Agric.104(1): 500–507. 10.1002/jsfa.12954
Huang, Y., Song, Y., Chen, F., Jiang, Z., Che, Z., Yang, X., and Chen, X. 2021. Simultaneous determination of eight biogenic amines in the traditional Chinese condiment Pixian Douban using UHPLC–MS/MS. Food Chem. 353: 129423. 10.1016/jfoodchem.2021.129423
Hurtado, A., Reguant, C., Bordons, A., and Rozès, N. 2012. Lactic acid bacteria from fermented table olives. Food Microbiol. 31(1): 1–8. 10.1016/j.fm.2012.01.006
International Olive Council (IOOC). 2004. Trade Standard Applying to Table Olives. International Olive Council, Madrid, Spain. https://www.internationaloliveoil.org/wp-content/uploads/2019/11/COI-OT-NC1-2004-Eng.pdf (Accessed on: 18 February 2025).
Jeon, A.R., Lee, J.H., and Mah, J.H. 2018. Biogenic amine formation and bacterial contribution in Cheonggukjang, a Korean traditional fermented soybean food. Food Sci Technol (LWT). 92: 282–289. 10.1016/j.lwt.2018.02.047
Kalinowska, K., and Tobiszewski, M. 2023. Green, simple analytical method for total biogenic amines content determination in wine using spectrophotometry. Food Chem. 402: 134457. 10.1016/j.foodchem.2022.134457
Kim, S.Y., Dang, Y.M., and Ha, J.H. 2022. Effect of various seasoning ingredients on the accumulation of biogenic amines in kimchi during fermentation. Food Chem. 380: 132214. 10.1016/j.foodchem.2022.132214
KučeroVá, K., SVobodoVá, H., Tůma, Š., Ondráčková, I., and Plocková, M. 2009. Production of biogenic amines by enterococci. Czech J Food Sci. 27(Special Issue 2): 50–55. 10.17221/673-CJFS
Lauková, A., Kandričáková, A., Buňková, L., Pleva, P., and Ščerbová, J. 2017. Sensitivity to enterocins of biogenic amine-producing faecal Enterococci from ostriches and pheasants. Probiotics Antimicrob. Proteins 9: 483–491. 10.1007/s12602-017-9272-z
Lázaro, C.A., Conte-Júnior, C.A., Canto, A.C., Monteiro, M.L.G., Costa-Lima, B., da Cruz, A.G., and Franco, R.M. 2015. Biogenic amines as bacterial quality indicators in different poultry meat species. Food Sci Technol (LWT). 60(1): 15–21. 10.1016/j.lwt.2014.09.025
Lee, J.H., Jin, Y.H., Lee, J.H., Park, Y.K., and Mah, J.H. 2024. Determination of biogenic amine-producing lactic acid bacteria in kimchi varieties through in vitro analysis and low temperature fermentation. Food Sci Biotechnol. 33: 2301-2312. 10.1007/s10068-024-01627-8
Li, B., and Lu, S. 2020. The importance of amine-degrading enzymes on the biogenic amine degradation in fermented foods: a review. Process Biochem. 99: 331–339. 10.1016/j.procbio.2020.09.012
Li, Y., Yan, T., Yin, L., Cheng, Y., and Jia, X. 2022. Isolation and identification of tyramine-producing bacteria and their biogenic amines formation during fermentation of sufu. Cell Mol Biol. 68(1): 75–88. 10.14715/cmb/2022.68.1.11
Liu, F., Du, L., Xu, W., Wang, D., Zhang, M., Zhu, Y., and Xu, W. 2013. Production of tyramine by Enterococcus faecalis strains in water-boiled salted duck. J Food Prot. 76(5): 854–859. 10.4315/0362-028X.JFP-12-487
Luo, Q., Shi, R., Gong, P., Liu, Y., Chen, W., and Wang, C. 2022. Biogenic amines in Huangjiu (Chinese rice wine): formation, hazard, detection, and reduction. Food Sci Technol (LWT). 168: 113952. 10.1016/j.lwt.2022.113952
Mah, J.H., Park, Y.K., Jin, Y.H., Lee, J.H., and Hwang, H.J. 2019. Bacterial production and control of biogenic amines in Asian fermented soybean foods. Foods 8(2): 85. 10.3390/foods8020085
Maijala, R.L. 1993. Formation of histamine and tyramine by some lactic acid bacteria in MRS-broth and modified decarboxylation agar. Lett Appl Microbiol. 17(1): 40–43. 10.1111/j.1472-765X.1993.tb01431.x
M’hir, S., Minervini, F., Di Cagno, R., Chammem, N., and Hamdi, M. 2012. Technological, functional and safety aspects of Enterococci in fermented vegetable products: a mini-review. Ann Microbiol. 62: 469–481. 10.1007/s13213-011-0363-x
Molaei, R., Tajik, H., and Moradi, M. 2019. Magnetic solid-phase extraction based on mesoporous silica-coated iron oxide nanoparticles for simultaneous determination of biogenic amines in an Iranian traditional dairy product Kashk. Food Control. 101: 1–8. 10.1016/j.foodcont.2019.02.011
Moniente, M., Botello-Morte, L., García-Gonzalo, D., Pagán, R., and Ontañón, I. 2022. Analytical strategies for the determination of biogenic amines in dairy products. Compr Rev Food Sci Food Saf. 21(4): 3612–3646. 10.1111/1541-4337.12980
Mounir, M., Hammoucha, J., Taleb, O., Afechtal, M., Hamouda, A., and Alaoui, M.I. 2021. Inoculation with acetic acid bacteria improves the quality of natural green table olives. Grasas Aceites 72(2): 407–407. 10.3989/gya.1259192
Mourad, K., and Nour-Eddine, K. 2006. Microbiological study of naturally fermented Algerian green olives: isolation and identification of lactic acid bacteria and yeasts along with the effects of brine solutions obtained at the end of olive fermentation on Lactobacillus plantarum. Grasas Aceites 57(3): 292–300. 10.3989/gya.2006.v57.i3.51
Müller, D.G., Oreste, E.Q., Heinemann, M.G., Dias, D., and Kessler, F. 2022. Biogenic amine sensors and its building materials: a review. Eur Polym J. 175: 111221. 10.1016/j.eurpolymj.2022.111221
Ovalle-Marmolejo, X.Y., Redondo-Solano, M., Granados-Chinchilla, F., Miranda-Castilleja, D.E., and Arvizu-Medrano, S.M. 2023. Effect of stress factors on the production of biogenic amines by lactic acid bacteria isolated from fermented Mexican foods (cheese and beer). Food Control 146: 109553. 10.1016/j.foodcont.2022.109553
Pereira, C.I., Matos, D., San Romão, M.V., and Barreto Crespo, M.T. 2009. Dual role for the tyrosine decarboxylation pathway in Enterococcus faecium E17: response to an acid challenge and generation of a proton motive force. Appl Environ Microbiol. 75(2): 345–352. 10.1128/AEM.01958-08
Portilha-Cunha, M.F., Macedo, A.C., and Malcata, F.X. 2020. A review on adventitious lactic acid bacteria from table olives. Foods 9(7): 948. 10.3390/foods9070948
Rehaiem, A., Fhoula, I., Slim, A.F., Boubaker, I.B.B., Chihi, A.B., and Ouzari, H.I. 2016. Prevalence, acquired antibiotic resistance and bacteriocin production of Enterococcus spp. isolated from Tunisian fermented food products. Food Control 63: 259–266. 10.1016/j.foodcont.2015.11.034
Sang, X., Li, K., Zhu, Y., Ma, X., Hao, H., Bi, J., and Hou, H. 2020. The impact of microbial diversity on biogenic amines formation in grasshopper sub shrimp paste during the fermentation. Front Microbiol. 11: 782. 10.3389/fmicb.2020.00782
Shalaby, A.R., Anwar, M.M., Sallam, E.M., and Emam, W.H. 2016. Quality and safety of irradiated food regarding biogenic amines: RAS cheese. Int J Food Sci Technol. 51(4): 1048–1054. 10.1111/ijfs.13058
Silva, I.P., Dias, L.G., da Silva, M.O., Machado, C.S., Paula, V.M.B., Evangelista-Barreto, N.S., and Estevinho, L.M. 2020. Detection of biogenic amines in mead of social bee. Food Sci Technol (LWT). 121: 108969. 10.1016/j.lwt.2019.108969
Sun, L., Guo, W., Zhai, Y., Zhao, L., Liu, T., Yang, L., and Duan, Y. 2023. Screening and the ability of biogenic amine-degrading strains from traditional meat products in Inner Mongolia. Food Sci Technol (LWT). 176: 114533. 10.1016/j.lwt.2023.114533
Tıraş, Z.E., and Yıldırım, H.K. 2021. Application of mixed starter culture for table olive production. Grasas Aceites 72(2): 405–405. 10.3989/gya.0220201
Tufariello, M., Anglana, C., Crupi, P., Virtuosi, I., Fiume, P., Di Terlizzi, B., and Bleve, G. 2019. Efficacy of yeast starters to drive and improve Picual, Manzanilla and Kalamàta table olive fermentation. J Sci Food Agric. 99(5): 2504–2512. 10.1002/jsfa.9460
Turna, N.S., Chung, R., and Mclntyre, L. 2024. A review of biogenic amines in fermented foods: occurrence and health effects. Heliyon 10: e24501. 10.1016/j.heliyon.2024.e24501
Vesković-Moračanin, S., Stefanović, S., Borović, B., Nastasijevic, I., Milijasevic, M., Stojanova, M., and Đukić, D. 2022. Assessment of biogenic amine production by lactic acid bacteria isolated from Serbian traditionally fermented foods. Acta Agric Serb. 27(53): 49‒55. 10.5937/AASer2253049V
Vinci, G., and Maddaloni, L. 2020. Biogenic amines in alcohol-free beverages. Beverages 6(1): 17. 10.3390/beverages6010017
Yalçınkaya, S., and Kılıç, G.B. 2019. Isolation, identification and determination of technological properties of the halophilic lactic acid bacteria isolated from table olives. J Food Sci Technol. 56(4): 2027–2037. 10.1007/s13197-019-03679-9
Yilmaz, N. 2024. Quantitative analysis of biogenic amine production of different lactic acid bacteria isolated from ready-to-eat packaged fish products. Vet Res Forum. 15(10): 537–543. 10.30466/vrf.2024.2024103.4193
Yilmaz, N., Özogul, F., Moradi, M., Fadiloglu, E.E., Šimat, V., and Rocha, J.M. 2022. Reduction of biogenic amines formation by foodborne pathogens using postbiotics in lysine-decarboxylase broth. J Biotechnol. 358: 118–127. 10.1016/j.jbiotec.2022.09.003
Zdolec, N., Mikuš, T., and Kiš, M. 2022. Lactic acid bacteria in meat fermentation: dry sausage safety and quality. In: Ray, R.C., Paramithiotis, S., Azevedo, V.A.deC., and Montet, D. (Eds.) Applied Biotechnology Reviews, Lactic Acid Bacteria in Food Biotechnology. Elsevier, Amsterdam, the Netherlands, Chap. 8, pp. 145–159. ISBN 9780323898751. 10.1016/B978-0-323-89875-1.00007-9
Zhang, Y., Shan, B., Gong, J., and Hu, Y. 2022. Mechanism of biogenic amine synthesis of Enterococcus faecium isolated from Sanchun ham. Food Sci Nutr. 10(6): 2036–2049. 10.1002/fsn3.2820