
Download

PAPER
Proximate analysis of lipid composition in Moroccan truffles and desert truffles
Fatima Henkrar1*, Lahsen Khabar2*
1Plant Biotechnology and Physiology Laboratory, Faculty of Sciences, University Mohammed V-Rabat, Morocco;
2Botanical, Mycology and Environment Laboratory, Faculty of Sciences, University Mohammed V-Rabat, Morocco
Abstract
Lipid composition in truffle is essential for nutraceutical and medicinal purposes. Currently, there is no data regarding the lipid content in Moroccan truffles. Therefore, we determined the fatty acid and sterol composition of six Moroccan truffles and desert truffles. The gas chromatography analysis revealed the predominance of fatty palmitic, oleic and linoleic acids. The prominent sterol components were brassicasterol and ergosterol. Besides, the sterol analysis discriminated between the Tuber and Terfezia truffles. These differences seem to be exploitable at a taxonomic level. This is a preliminary report disclosing the fatty acid and sterol composition of Moroccan truffles, indicating the potential use of lipids analysis, especially sterol analysis, as biomarker for truffles distinction.
Key words: desert truffles, discrimination, fatty acid, gas chromatography, Moroccan truffle, sterol
*Corresponding Authors: Fatima Henkrar, Plant Biotechnology and Physiology Laboratory, Faculty of Sciences, University Mohammed V-Rabat, Morocco. Email: [email protected]; Lahsen Khabar, Botanical, Mycology and Environment Laboratory, Faculty of Sciences, University Mohammed V-Rabat, Morocco. Email: [email protected]
Received: 18 March 2022; Accepted: 17 May 2022; Published: 10 June 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
The truffles are edible ascocarp of hypogenous ascomycetes fungi that grow underground (Khabar et al., 2001; Lee et al., 2020). The term ‘desert truffles’ is used to describe truffles growing particularly in arid and semi-arid areas (Khabar et al., 2001; Morte et al., 2021), such as Morocco, Algeria, Tunisia, Egypt, South Africa, Saudi Arabia, Iraq, Syria and Kuwait (Khabar, 2014; Lee et al., 2020). The genera found abundantly in those areas are Terfezia and Tirmania. Besides, other genera exist as well, namely Delastria and Picoa (Khabar, 2014). In Mediterranean countries, especially in North Africa, the truffles are harvested in abundance and known as ‘Terfass,’ also called ‘Kame,’ ‘Kholassi,’ ‘Zoubaïdi,’ ‘Truffles of the deserts’ and ‘Truffles of the sands’ because of their development in sandy soil (Khabar, 2014).
The determination of lipid composition in truffles is essential both for lipid analysis as well as for nutraceutical and medicinal purposes. The truffles contain only 4–9% by dry weight of total lipids (Tang et al., 2011). Fatty acids and phytosterols are the main lipid compounds in truffle fruiting bodies, which are well known for their potential human benefits. Several studies through global chromatographic analysis demonstrated that desert truffles are rich in fatty acids, both saturated and unsaturated that have many positive effects on health (Akyüz, 2013; Al-Shabibi et al., 1982; Bokhary et al., 1989; Doğan and Aydın, 2013; Veeraraghavan et al., 2021). For example, in Terfezia boudieri from Tunisia, Hamza et al. (2016) reported that essential fatty acids, like linoleic and oleic acids, account for 76% of the total fat content. Linoleic acid or omega-6 is an essential fatty acid and one of the most aromatic compounds in most truffle species (Lee et al., 2020), which has protective and antioxidative effects beneficial for human health (Sokoła-Wysoczańska et al., 2018). While, oleic acid, a bioactive compound, has the aptitude in reducing cholesterol levels (Lee et al., 2020). Another example of Terfezia claveryi from Saudi Arabia, which is closely related to T. boudieri (Dahham et al., 2018), was found to have arachidic, myristic, palmitic, behenic, pentadecanoic, stearic, heneicosanoic, nonadecanoic and margaric acids as saturated fatty acids along with unsaturated fatty acids (palmitoleic, oleic, erucic and linoleic) (Bokhary et al., 1989). The lipid composition of desert truffles depends highly on the species as well as growing environments. For instance, T. boudieri from Saudi Arabia was rich in pentadecanoic, margaric, stearic and arachidic acids (Bokhary et al., 1989). Whereas, the same species from Turkey contained mainly oleic, linoleic, linolenic, palmitic, palmitoleic, stearic and behenic acids (Akyüz, 2013). The most identified phytosterols in truffle reports were brassicasterol and ergosterol. Harki et al. (1996) reported that the prominent components identified in Tuber melanosporum were ergosterol and brassicasterol, accounting for about 90% of total sterols. As well, the major sterol components in the Tuber ascocarps were brassicasterol and ergosterol, accounting for about 17–64% and 25–67% of total sterols, respectively (Tang et al., 2012). In Terfezia truffles, brassicasterol levels were 98% of the total sterols, while ergosterol was present in lower amounts (Tejedor-Calvo et al., 2021). Other phytosterols such as beta-sitosterols, stigmasterol and campestanol were also present in low contents (Dahham et al., 2018).
The six species included in this study were natives of Morocco. Terfezia arenaria, commonly called ‘Pink Terfess of Mamora,’ was harvested from acidic soil, in semi-arid climate under Helianthemum guttatum. It is an appreciated edible fungus, detected by the ‘mark’ method (Khabar, 2014) unlike Delastria rosea, known as ‘Bitter Terfess of Taida’ due to its bitter flavour. It was collected under Pinus pinaster var. atlantica and Pinus halepensis in Mamora forest between November and December (Khabar, 2014). Similarly, Tuber oligospermum was also collected under P. pinaster var. atlantica in Mamora forest, starting from December until April. The ascocarp of T. boudieri originated from Bouaarfa region, collected from limestone soil, under arid and sub-Saharan climate, and Terfezia leptoderma was obtained from Mamora forest from the acidic soil under H. guttatum at the beginn-ing of February until May. Tuber asa, commonly called ‘Terfass male of Terfass’ because of its hard consistency, collected as well under H. guttatum on acidic soil of Mamora forest, towards the end of February at the same time as T. leptoderma (Khabar, 2014). The aims of this work were 1) to determine the lipid profile of the six Moroccan truffle species, 2) to determine the relation between the genus, species and lipid composition of truffles, and 3) to determine whether the lipid profile can be used as a tool for species or genus distinction.
Materials and methods
Fungus material
Six Moroccan truffle species were used in this experiment. The ascocarps of different species (Table 1) were collected directly from their natural environments and transported to the laboratory. Under the fume hood, the samples were surface sterilised with ethanol, peeled and then fragmented by hand. Several pieces were taken from the glebe and stored in pillboxes at −64°C. Alternatively, other samples were sun-dried for 2 months before being stored at −64°C.
Table 1. The name and location of six Moroccan truffle species used in this study.
| Species name | Location |
|---|---|
| Terfezialeptoderma (1) | Mamora Forest under Helianthemum |
| T.leptoderma (2) | Mamora Forest under Pinus pinaster |
| Terfeziaarenaria | Mamora Forest |
| Terfeziaboudieri | Bouaarfa |
| Tuber asa | Mamora Forest |
| Tuber oligospermum | Mamora Forest under P. pinaster |
| Delastriarosea | Mamora Forest under P. pinaster |
The different steps of extraction, separation and analy-sis of lipids were released at the Laboratory of Myco-logy, Phytopathology and Environment of the Littoral France University, following the method of Fontaine et al. (2001).
Extraction of total lipids
The extraction was performed with approximately 20–40 mg of freeze-dried material (pieces of truffle glebe). The solvent used for extraction was a mixture of dichloromethane and methanol (2:1 v/v) with 0.05% BHT (Butylated hydroxytoluene; Sigma) as antioxidant. The freeze-dried fungal material was first ground in 40 ml of the solvent using ultra-turrax homogenizer. The first extractions were performed in the dark to preserve the ergosterol, a photosensitive sterol. The extraction of total lipids was carried out under reflux (1 h at 70°C) with some pieces of pumice stone. After filtration, the lipid extract was recovered under nitrogen blowdown and rotary evaporator at 60°C. This step was repeated thrice.
Separation of fatty acids and total sterols by saponification
The crude lipid extract was used to separate fatty acids and total sterols by saponification. The crude extract was saponified under reflux (1 h at 90°C) in 2 ml of 6% (w/v) methanolic potash and some pieces of pumice. After cooling, two successive cold extractions with hexane were performed. The first extraction permitted the recovery of unsaponifiable fraction (sterols), while the second one enabled the retrieval of saponifiable fraction (fatty acid). To perform the first cold extraction, one volume of distilled water was added to the cooled saponified extract, followed by six volumes of hexane. This mixture was then vigorously stirred for 1 min with a vortex. After decantation, the organic phase (upper layer), which contains the unsaponifiable elements, was taken out and dehydrated with anhydrous sodium sulfate. This step was repeated three times, and the recovered extract was concentrated in a rotary evaporator at 50°C. For the fatty acid recuperation, the aqueous phase was acidified to pH 1 with 1 M HCl to liberate them from their saline combination. Afterwards, the acidified phase was extracted by performing three extractions with hexane. The concentration of these extracts was done under nitrogen.
Fatty acid analysis
The fatty acids were solubilise in 1 ml boron trifluoride–methanol (14% w/v). The methylation reaction was carried out for 2 min at 90°C in a water bath and then stopped by immersing the tubes in ice. After addition of 1 ml of distilled water and 2 ml of hexane, the tubes were vortexed for 30 s. The organic phase (upper phase) was taken out and dehydrated by adding anhydrous sodium sulfate. This step was repeated thrice. The methylated fatty acids were purified on silica gel of TLC (20 × 20 cm, type Silicagel F 254, Merck) with a solvent system composed of diethyl ether/hexane (10/90; v/v). The fatty acid spots were detected by primuline and eluted in about 1 ml of dichloromethane. Thereafter, the fatty acids were immediately taken up in 25–100 µl of hexane and injected into the gas chromatograph. Fatty acids were identified by comparing their relative retention times with internal standard such as methyl C 17:0 (methyl margarate) as well as other known standards (Alltech).
Sterol analysis
To obtain a better separation in gas chromatography, sterols were acetylated either for 12 h at room temperature or 2 h at 60°C by the mixture of toluene/acetic anhydride/pyridine (1/2/1; v/v/v). Sterol acetates were purified on silica gel thin layer with dichloromethane as migration solvent. Cholesterol acetate (1 mg/ml) was used as a control to localise acetylated products after spraying with 0.01% (w/v) primuline solution. The acetylated sterols were taken up in 25–100 µl of hexane and injected into the chromatograph. The sterol acetates were identified by comparing their relative retention times against an internal standard, cholesterol in alcohol form (non-acetylated) along with brassicasterol and other known acetylated standards.
Statistical analysis
The data were analysed using R studio software. The means and standard deviations were calculated. The pairwise comparisons among means were performed using two-way ANOVA and Tukey HSD test. To indicate significant differences, we used the multcompLetters4() function from the multcompView package.
Results and discussions
The objective of this work was to define the nature and proportion of fatty acids and sterols in six species of Moroccan truffles and to determine if this lipid profile could be used as a classification tool to discriminate between species or genus. This study was conducted for the first time on Moroccan truffles, disclosing distinctly the fatty acid and sterol components of truffles and desert truffles grown in Morocco.
Fatty acid composition
The fatty acid composition of Moroccan truffles has not been reported previously, and studies on fatty acid content of other truffles are scarce. The first and most reported studies on fatty acid composition were focused on Tirmania pinoyi, Tirmania nivea, T. boudieri, T. claveryi and Picoa lefebvrei from Saudi Arabia (Bokhary et al., 1989; Bokhary and Parvez, 1995) and T. claveryi from Iraq (Al-Shabibi et al., 1982). Recently, other studies appeared on fatty acid composition of T. boudieri from Iraq (Dahham et al., 2018), T. boudieri from Turkey (Hamza et al., 2016), T. claveryi and Picoa juniperi from Spain (Murcia et al., 2003) as well as T. nivea from Libya (Shah et al., 2020).
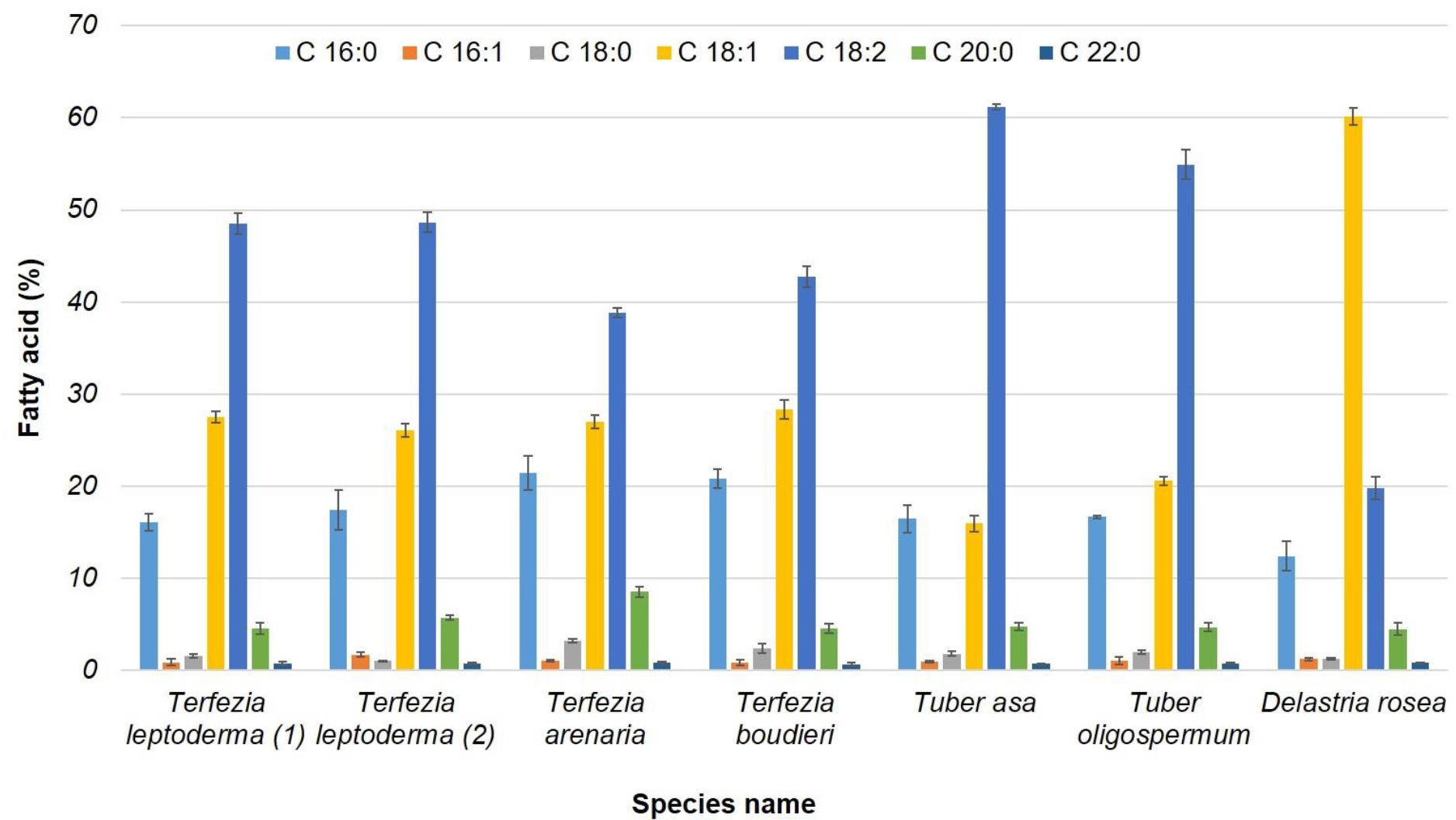
The chromatographic analysis results for the identification of fatty acids compositions are presented in (Table 2 and Figure 1). Seven fatty acids were detected in the six truffles species used in this experiment; four saturated fatty acids [palmitic (C16:0), stearic (C18:0), arachidic (C20:0) and behenic (C22:0)], and three unsaturated fatty acids [palmitoleic (C16:1), oleic (C18:1) and linoleic (C18:2)]. Bokhary et al. (1989) reported that palmitic, stearic, oleic and linoleic acids were predominant in T. nivea and T. boudieri which originated from Saudi Arabia. As well, the Turkish T. boudieri was also rich in behenic, palmitic, palmitoleic, stearic, oleic, linoleic and linolenic acids (Akyüz, 2013), which agree with our results and particularly with T. boudieri. Furthermore, a recent study on fatty acid composition in Tuber maculatum, Tuber aestivum/uncinatum, Tuber borchii, T. melanosporum and T. nivea revealed the dominance of palmitic, stearic, oleic and linoleic acids followed by traces of polyunsaturated fatty acids (Shah et al., 2020).
Table 2. Fatty acid composition of the six Moroccan truffle species through gas–liquid chromatography analysis (percentage of dry weight of the lipid fraction).
| Species | C 16:0 | C 16:1 | C 18:0 | C 18:1 | C 18:2 | C 20:0 | C 22:0 |
|---|---|---|---|---|---|---|---|
| Terfezialeptoderma (1) | 16.080 ± 0.936a | 0.913 ± 0.372a | 1.600 ± 0.185a | 27.530 ± 0.598a | 48.513 ± 1.179a | 4.570 ± 0.589a | 0.790 ± 0.147a |
| T.leptoderma (2) | 17.456 ± 2.167a | 1.733 ± 0.221a | 1.026 ± 0.047a | 26.106 ± 0.740a | 48.633 ± 1.075a | 5.753 ± 0.247a | 0.803 ± 0.100a |
| Terfeziaarenaria | 21.470 ± 1.822b | 1.066 ± 0.133a | 3.253 ± 0.206a | 27.020 ± 0.727a | 38.803 ± 0.525b | 8.563 ± 0.577b | 0.900 ± 0.095a |
| Terfeziaboudieri | 20.823 ± 1.019b | 0.830 ± 0.303a | 2.433 ± 0.508a | 28.333 ± 1.028a | 42.770 ± 1.127c | 4.580 ± 0.545a | 0.710 ± 0.156a |
| Tuber asa | 16.470 ± 1.483a | 0.940 ± 0.096a | 1.810 ± 0.259a | 15.983 ± 0.879b | 61.186 ± 0.315d | 4.773 ± 0.396a | 0.740 ± 0.070a |
| Tuber oligospermum | 16.670 ± 0.138a | 1.090 ± 0.389a | 2.023 ± 0.200a | 20.593 ± 0.450c | 54.926 ± 1.601e | 4.720 ± 0.500a | 0.780 ± 0.105a |
| Delastria rosea | 12.420 ± 1.574c | 1.230 ± 0.166a | 1.263 ± 0.112a | 60.160 ± 0.916d | 19.836 ± 1.237f | 4.503 ± 0.678a | 0.830 ± 0.052a |
Data shown as mean ± standard deviation (n = 3). Different superscript letters in the same column indicate a statistically significant difference (P < 0.05).
Figure 1. Fatty acid composition of the six Moroccan truffle species used in this study.
The main fatty acids detected were palmitic, oleic and linoleic acids. The other fatty acids were also present but at lower levels. Similar findings were also reported in various species of Terfezia and Tuber (Hamza et al., 2016; Tejedor-Calvo et al., 2021). Our results demonstrated that the rate of palmitic acid (C16:0) is appreciably equal in all the species studied. We could also notice that the linoleic acid level was generally higher compared to the oleic acid (C18:1) level in all the species studied except for D. rosea where the opposite was true; the level of oleic acid was higher than linoleic acid. The same results were also reported by Hamza et al. (2016). The T. boudieri was characterised by its higher content of linoleic acid (54.10%) compared to oleic and palmitic acids that represented 22 and 20.40%, respectively (Hamza et al., 2016). Linoleic acid level was considerably high in Tuber compared to Terfezia species. The rate of oleic acid was, on the other hand, slightly lower in Tuber genus compared to Terfezia genus. This remark goes with Tejedor-Calvo et al. (2021), who reported that linoleic acid content in Tuber brumal and T. melanosporum reached 78.3 and 61.12%, respectively, compared to T. leptoderma and T. arenaria which noticed only 51.3 and 30.9%, respectively. The fatty acid results of Moroccan truffles and desert truffles proved their richness in unsaturated and healthy fatty acids such as linoleic acid, suggesting their equivalent culinary value compared to European truffles.
Regarding fatty acid discrimination, it seems that these criteria could not differentiate clearly between the species of the two genera of Tuber and Terfezia. Indeed, the ratio of linoleic acid or oleic acid was approximately equal between the different species of the two genera. Nevertheless, the fatty acid composition could distinguish between Delastria and other two genera.
Sterol composition
The sterol composition of Moroccan truffles has never been reported before. The first report on sterol was that by Weete et al. (1985), which mentioned about both Terfezia and Tuber genera. Further, other studies principally on Tuber species including T. melanosporum (Harki et al., 1996; Sancholle et al., 1988), Tuber magnatum, T. melanosporum, T. aestivum, Tuber albidum and Tuber indicum were released (Sommer et al., 2020).
Four sterols (brassicasterol, ergosterol and lanosterol) were identified in the ascocarps of the examined truffle species (Table 3 and Figure 2). Furthermore, the main sterols were ergosterol and brassicasterol with the highest percentage in all the examined species. Similar results were disclosed in black truffle, where the sterol composition of T. melanosporum was analysed and ergosterol along with brassicasterol were identified as the major components (90%) (Harki et al., 1996). As well, the examination of Tuber sinense, T. aestivum, T. indicum, Tuber himalayense and T. borchii revealed the predominance of brassicasterol and ergosterol, with 17–64% and 25–67% of total sterols, respectively (Tang et al., 2012).
Table 3. Sterol composition of the six Moroccan truffle species through gel permeation chromatography analysis.
| Sterols | Brassicasterols | Ergosterols | Lanosterol (1.31) | n.i. (1.42) |
|---|---|---|---|---|
| Terfezialeptoderma (1) | 96.870 ± 1.260a | 3.016 ± 0.621a | 0 ± 0a | n.d. |
| T.leptoderma (2) | 92.410 ± 2.606a | 8.153 ± 0.143a | 0 ± 0a | n.d. |
| Terfeziaarenaria | 96.833 ± 1.045a | 0 ± 0b | 2.003 ± 0.532a | n.d. |
| Terfeziaboudieri | 97.190 ± 2.416a | 0 ± 0b | 3.110 ± 0.298a | n.d. |
| Tuber asa | 40.526 ± 2.377b | 23.196 ± 0.718c | 17.470 ± 0.856b | 12.220 ± 2.186b |
| Tuber oligospermum | 46.006 ± 2.001c | 21.470 ± 1.990c | 16.223 ± 4.441b | 17.286 ± 1.997b |
| Delastria rosea | 21.343 ± 1.770d | 42.9633 ± 0.621d | 22.826 ± 3.471c | 12.886 ± 1.806b |
Data shown as mean ± standard deviation (n = 3). Lanosterol and n.i. compounds are reported with their retention time (in minutes) between brackets. Different superscript letters in the same column indicate a statistically significant difference (P < 0.05). n.i., not identified; n.d., not detected.
Figure 2. Sterol composition of the six Moroccan truffle species.
Besides, the ratio of ergosterol to brassicasterol changes according to the genera studied. In Terfezia species, brassicasterol was the main sterol identified, accounting for 92–97% of the total sterols, affirming the results of Weete et al. (1985), who reported that brassicasterol levels were 98% of total sterols in Terfezia truffles, while ergosterol was registered at very low amounts (0–8%). On the other hand, in Tuber species (Tuber asa and Tuber oligospermum) and D. rosea, the ergosterol represented a considerable amount compared to Terfezia species, accounting for 23, 21 and 43% of the sterols, respectively. These species also contained 40, 46 and 21% of brassicasterol, respectively. A recent study by Tejedor-Calvo et al. (2022) demonstrated that ergosterol and brassicasterol were the two main sterols in T. claveryi and T. aestivum ascocarps, with differences in ergosterol to brassicasterol ratio depending on the ascocarp genus. Lanosterol was also detected in Tuber species, as well as in D. rosea, in considerable quantities, accounting for 16 and 23% of the sterols, respectively. While in Terfezia, they were either totally absent or present in very small quantity (approximately 2–3%). The high amount of brassicasterol in Terfezia will increase the quality interest of the Moroccan genera, knowing that brassicasterol has several health benefits, such as antioxidative activity and anti-infective properties.
Finally, Terfezia genus was distinguished by the high brassicasterol content, while the Tuber genera and Delastria were characterised by the equivalent amount of ergosterol and brassicasterol. Hence, sterol analysis proved their importance to highlight differences between species and to separate the Tuber from Terfezia truffles collected in Morocco. These differences seem to be exploitable at the taxonomic level.
Conclusion
The lipid composition and concentration were highly influenced by truffle speciation and their growing area. This was the first report of lipid composition of Moroccan truffles, divulging the fatty acid and sterol compositions of six species of truffles and desert truffles and demonstrating the richness of Moroccan truffles in essential unsaturated fatty acid, such as linoleic acid. There was a slight difference between Tuber and Terfezia species in fatty acid component, which is not sufficient to differentiate between them. However, sterol analysis distinguished between these two genera. Hence, a comparison of their sterol composition with reported data seems to be plausible for Tuber and Terfezia distinction. Finally, a deeper study on other nutrient compounds and bioactive molecules of Moroccan truffles as well as their antioxidant evaluation is predetermined to improve their edible and culinary interest through their health benefits.
REFERENCES
Akyüz, M., 2013. Nutritive value, flavonoid content and radical scavenging activity of the truffle (Terfezia boudieri Chatin). Journal of Soil Science and Plant Nutrition. 13:143–151.
Al-Shabibi, M.M.A., Toma, S.J. and Haddad, B.A., 1982. Studies on Iraqi truffles. I. Proximate analysis and characterization of lipids. Canadian Institute of Food Technology Journal. 15:200–202. 10.4067/S0718-95162013005000013
Bokhary, H.A. and Parvez, S., 1995. Studies on the chemical composition of the Ascomycete fungus Phaeangium lefebvrei Pat. Journal of King Saud University. 7:215–224.
Bokhary, H.A., Suleiman, A.A. and Basalah, M.O., 1989. The fatty acid components of the desert truffle ‘Al Kamah’ of Saudi Arabia. Journal of Food Protection. 52(9):668–669. 10.4315/0362-028X-52.9.668
Dahham, S.S., Al-Rawi, S.S., Ibrahim, A.H., Aman, S.A.M. and Amin, M.S.A.M., 2018. Antioxidant, anticancer, apoptosis properties and chemical composition of black truffle Terfezia claveryi. Saudi Journal of Biological Sciences. 25:1524–1534. 10.1016/j.sjbs.2016.01.031
Doğan, H.H. and Aydın, S., 2013. Determination of antimicrobial effect, antioxidant activity and phenolic contents of desert truffle in Turkey. African Journal of Traditional, Complementary and Alternative Medicines. 10:52–58.
Fontaine, J., Grandmougin-Ferjani, A., Hartmann, M.A. and Sancholle, M., 2001. Sterol biosynthesis by the arbuscular mycorrhizal fungus Glomus intraradices. Lipids. 36:1357–1363. 10.1007/s11745-001-0852-z
Hamza, A., Zouari, N., Zouari, S., Jdir, H., Zaidi, S., Gtari, M. and Neffati, M., 2016. Nutraceutical potential, antioxidant and antibacterial activities of Terfezia boudieri Chatin, a wild edible desert truffle from Tunisia arid zone. Arabian Journal of Chemistry. 9:383–389. 10.1016/j.arabjc.2013.06.015
Harki, E., Klaebe, A., Talou, T. and Dargent, R., 1996. Identification and quantification of Tuber melanosporum Vitt. sterols. Steroids. 61:609–612. 10.1016/s0039-128x(96)00121-3
Khabar, L., 2014. Mediterranean basin: North Africa. Ch.10. In: Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y. and Morte, A. (eds.) Desert truffles: phylogeny, physiology, distribution and domestication. Springer, Berlin, Heidelberg, pp. 143–158.
Khabar, L., Najim, L., Janex-Favre, M. and Paraguey-Leduc, A., 2001. Contribution à l’étude de la flore mycologique du Maroc. Les truffes marocaines. Bulletin de la Société mycologique de France. 117:213–229.
Lee, H., Nam, K., Zahra, Z. and Farooqi, M.Q.U., 2020. Potentials of truffles in nutritional and medicinal applications: a review. Fungal Biology Biotechnology. 7(9):1–17. 10.1186/s40694-020-00097-x
Morte, A., Kagan-Zur, V., Navarro-Ródenas, A. and Sitrit, Y., 2021. Cultivation of desert truffles—a crop suitable for arid and semi-arid zones. Agronomy. 11:1462. 10.3390/agronomy11081462
Murcia, M.A., Martínez-Tomé, M., Vera, A., Morte, A., Gutierrez, A., Honrubia, M. and Jiménez, A.M., 2003. Effect of industrial processing on desert truffles Terfezia claveryi Chatin and Picoa juniperi Vittadini): proximate composition and fatty acids. Journal of the Science of Food and Agriculture. 83:535–541. 10.1002/jsfa.1397
Sancholle, M., Weete, J.D., Kulifaj, M. and Montant, C., 1988. Changes in lipid composition during ascocarp development of the truffle, Tuber melanosporum. Mycologia. 80:900–903. 10.2307/3807580
Shah, N.N., Hokkanen, S., Pastinen O., Eljamil, A. and Shamekh, S., 2020. A study on the fatty acid composition of lipids in truffles selected from Europe and Africa. 3 Biotech. 10:415. 10.1007/s13205-020-02414-y
Sokoła-Wysoczańska, E., Wysoczański, T., Wagner, J, Czyż, K., Bodkowski, R., Lochyński, S. and Patkowska-Sokoła, B., 2018. Polyunsaturated fatty acids and their potential therapeutic role in cardiovascular system disorders—a review. Nutrients. 10(10): 1561. 10.3390/nu10101561
Sommer, K., Krauß, S. and Vetter, W., 2020. Differentiation of European and Chinese truffle (Tuber sp.) species by means of sterol fingerprints. Journal of Agricultural and Food Chemistry. 68(49):14393–14401. 10.1021/acs.jafc.0c06011
Tang, Y., Li, H.-M. and Tang, Y.-J., 2012. Comparison of sterol composition between Tuber fermentation mycelia and natural fruiting bodies. Food Chemistry. 132(3):1207–1213. 10.1016/j.foodchem.2011.11.077
Tang, Y., Li, Y.-Y., Li, H.-M., Wan, D.-J. and Tang, Y.-J., 2011. Comparison of lipid content and fatty acid composition between tuber fermentation mycelia and natural fruiting bodies. Journal of Agricultural and Food Chemistry. 59:4736–4742. 10.1021/jf200141s
Tejedor-Calvo, E., Amara, K., Reis, F.S., Barros, L., Martins, A., Calhelha, R.C., Venturini, M.E., Blanco, D., Redondo, D., Marco, P. and Ferreira, I.C.F.R., 2021. Chemical composition and evaluation of antioxidant, antimicrobial and antiproliferative activities of tuber and Terfezia truffles. Food Research International. 140:110071. 10.1016/j.foodres.2020.110071
Tejedor-Calvo, E., García-Barreda, S., Sánchez, S., Morte, A., Siles-Sánchez, M.D.N., Soler-Rivas, C., Santoyo, S. and Marco, P., 2022. Application of pressurized liquid extractions to obtain bioactive compounds from Tuber aestivum and Terfezia claveryi. Foods. 11(3):298. 10.3390/foods11030298
Veeraraghavan, V.P., Hussain, S., Balakrishna, J.P., Dhawale, L., Kullappan, M., Ambrose, J.M. and Mohan, S.K., 2021. A comprehensive and critical review on ethnopharmacological importance of desert truffles: Terfezia claveryi, Terfezia boudieri, and Tirmania nivea. Food Reviews International. 0:1–20. 10.1080/87559129.2021.1889581
Weete, J.D., Kulifaj, M., Montant, C., Nes, W.R. and Sancholle, M., 1985. Distribution of sterols in fungi. II. Brassicasterol in Tuber and Terfezia species. Canadian Journal of Microbiology. 31: 1127–1130. 10.1139/m85-212