
Download



REVIEW
A systematic review to introduce the most effective postbiotics derived from probiotics for aflatoxin detoxification in vitro
Setayesh Zamanpour1,2, Seyyed Mohammad Ali Noori3,4, Behdad Shokrollahi Yancheshmeh5,6, Asma Afshari1,2, Mohammad Hashemi1,2*
1Medical Toxicology Research Center, Mashhad University of Medical Sciences, Mashhad, Iran;
2Department of Nutrition, Faculty of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran;
3Toxicology Research Center, Medical Basic Sciences Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran;
4Department of Nutrition, School of Allied Medical Sciences, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran;
5Research Group for Food Production Engineering, National Food Institute, Technical University of Denmark, Lyngby, Denmark;
6Food Safety Research Center (Salt), Semnan University of Medical Sciences, Semnan, Iran
Abstract
The purpose of this study was to gather information about the percentage of aflatoxin decontamination by postbiotics and to find the most effective postbiotic. In this review article, studies were collected from databases. All the articles related to experimental studies were included in the study. It appeared that the postbiotics derived from Lentilactobacillus kefiri, Lentilactobacillus kefiri, Lacticaseibacillus rhamnosum, Lacticaseibacillus rhamnosum and Pediococcus pentosaceus probiotic bacteria were the most effective postbiotics with 97.22%, 95.27%, 86.2%, 81.4% and 91% inhibitory effect against aflatoxins B1, B2, G1, G2 and M1, respectively. Therefore, postbiotics could be used as safe anti-aflatoxin agents in food products.
Key words: aflatoxins, cell-free supernatant, killed probiotics, non-viable probiotics, postbiotics
*Corresponding Author: Mohammad Hashemi, Medical Toxicology Research Center, Mashhad University of Medical Sciences, Mashhad, Iran. Email: [email protected]
Received: 14 May 2023; Accepted: 1 September 2023; Published: 6 October 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Aspergillus flavus and Aspergillus parasiticus produce aflatoxins (AFs) which have mutagenic, carcinogenic, hepatotoxic, immunosuppressive and teratogenic effects in humans and animals, and are responsible for the loss of crops and animal feed (Hernandez-Mendoza et al., 2009a, 2009b). They are present in food products, such as corn, peanuts, beans, dried fruits and dairy products, including milk (Ambadoyiannis et al., 2004; Dalié et al., 2010). Owing to their high thermal stability, aflatoxins remain in raw food and are not destroyed by processing and cooking, and are trasferred to humans through food (Zain, 2011). Since mycotoxins have low molecular weight, they are promptly absorbed in the gastrointestinal tract (GIT) and can be discovered in blood and milk 15 min and 12 h after intake, respectively (Martins et al., 2001). More than 20 metabolites of the AF group are known, among which the most important ones are B1, B2, G1 and G2 with different levels of toxicity (Alshannaq and Yu 2017; Laciakova et al., 2008; Mahmood et al., 2018). These terms are derived through fluorescence under ultraviolet (UV) rays, blue (B) or green (G), and comparative chromatographic migration patterns through thin layer chromatography (TLC) (Lizárraga-Paulín et al., 2011; Vosough et al., 2013). AFM1 and AFM2 are hydroxylated metabolites of AFB1 and AFB2. Approximately 0.3–6.2% of AFB1 ingested by animals is metabolized to AFM1 (Figure 1) (Afshar et al., 2020; Battacone et al., 2003; Kumar et al., 2017; Oatley et al., 2000).
Figure 1. Sources of aflatoxin exposure and its flow in the food chain.
The order of acute and chronic toxicity of different aflatoxins is AFB1 > AFG1 > AFB2 > AFG2, where AFM1 and AFM2 are less toxic than their precursors (Ahmadi, 2020). Among these, AFB1 showed the highest toxicity for animals and humans (Dhanasekaran et al., 2011; Sklan et al., 2001; Yu et al., 2008). The International Agency for Research on Cancer (IARC) classified AFB1 and AFM1 as human carcinogens belonging to Group 1 and Group 2B, respectively (Caloni et al., 2006; Darwish et al., 2014; Marchese et al., 2018). The Codex Alimentarius specifies maximum limit of aflatoxins as 15 µg/kg for almonds, hazelnuts, Brazil nuts, peanuts and pistachio nuts for further processing, and 10 µg/kg for almonds, Brazil nuts, hazelnuts and pistachio nuts for direct consumption and dried figs, with a maximum limit of AFM1 as 0.5 µg/kg for milk. However, the maximum levels of aflatoxins in foods vary globally, depending on the type of product and also on the import/export regime (Food and Agriculture Organization (FAO), 2003; Pickova et al., 2021). There are various approaches to reduce or remove mycotoxins, including physical (thermal inactivation, radiation, magnetic carbon absorption), chemical (treatment with acid, alkali, or oxidizing agent), and biological (microorganisms) treatments (Peng et al., 2018). Among these methods, biological methods attracted a great deal of attention because of their high efficiency, low price, compatibility with the environment, and complete detoxification under favorable conditions (Adebo et al., 2017; Armando et al., 2012; Shetty and Jespersen, 2006). Among the biological methods, detoxification using postbiotics has gradually become the most appropriate biological detoxification method, because the use of microorganisms in food chains required a long process of national and international authorization (Zhou et al., 2017; Zychowski et al., 2013).
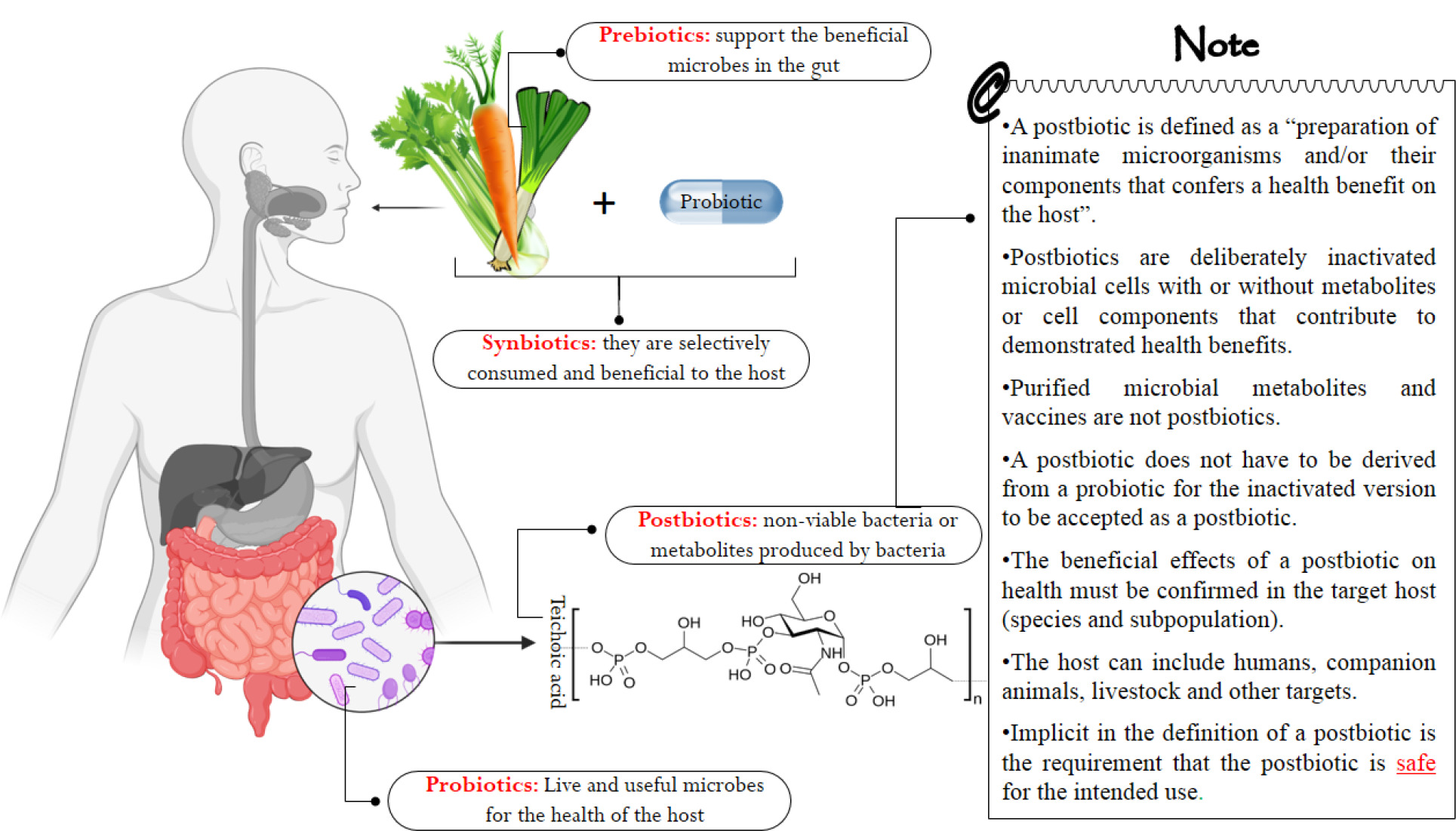
Some microorganisms, especially probiotic bacteria, have received special attention because of their various beneficial effects, such as an easy, controllable, and safe method without loss of nutritional value to remove aflatoxins (Mahmood et al., 2018). Along with the remarkable progress made in this field, the researchers got acquainted with a new concept of probiotics called postbiotics. The word “postbiotic” refers to bioactive compounds and does not match the definitions of prebiotics, synbiotics and probiotics. In mid-2021, the International Scientific Association for Probiotics and Prebiotics (ISAPP) provided a precise and well-considered definition of postbiotics, which is the “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” (Salminen et al., 2021). According to the above definition, the term postbiotic refers to a set of macro and micro molecules, such as inactive microorganisms (cells that are not alive), parts of a cell (surface layer proteins, endo- and -exo-polysaccharides, teichoic acid, muropeptides and cell-free supernatants [CFS]), or metabolites of a cell (bacteriocins, short-chain fatty acids [SCFA], organic acids and enzymes that are produced naturally by living probiotic cells during fermentation) (Figure 2) (Homayouni-Rad et al., 2021).
Figure 2. Definition of the family of probiotics, prebiotics, synbiotics and postbiotics (Vinderola et al., 2022).
Terms such as parapsychobiotics, paraprobiotics, non--viable probiotics, heat-killed probiotics, ghost probiotics, meta-biotics, cell-free supernatant, biogenic, tyndallized probiotics, bacterial lysates, and postbiotics are also used (Homayouni-Rad et al., 2021; Salminen et al., 2021), although term postbiotic is used more commonly.
Probiotics are mostly lactic acid bacteria (Walhe et al., 2022) and have many health benefits; however, because they are alive, they have limitations for use and treatment. Probiotic bacteria must maintain their viability even in inappropriate conditions, such as preparation of food, storage, processing, distribution as well as passing through the digestive system, so that they can perform their health-giving effects (Homayouni-Rad et al., 2020; Rad et al., 2020a). In spite of passing these challenges and survival, adverse reactions may occur, such as resistance gene transfer between other intestinal microbiota, different colonization patterns, presence of virulence factors in some probiotic species, preventing the colonization of other intestinal microbiota, meningitis, infectious endocarditis, urinary tract infections, and bacteremia, especially in infants, patients treated with antibiotics and have weak immunity (Abbasi et al., 2021a, 2021b; Abriouel et al., 2015; Kim et al., 2018; Liong 2008).
Postbiotics have overcome these adverse effects and appeared as a suitable alternative to probiotics. In recent years, it has been shown that the viability of a probiotic is not necessarily to benefit from its favorable effects (Piqué et al., 2019). It has been proven both in vivo and in vitro that the beneficial effects of probiotics and postbiotics include antibacterial, antiviral, antifungal, antioxidant, anti-proliferation, anti-mutation, anti--cancer, -anti-obesity, anti-diabetic, anti-blood pressure, and immune system booster (Moradi et al., 2021). In addition, postbiotics have unique features, such as safe origin, no toxic effect, low cost of preparation and transportation, and long shelf life (Homayouni-Rad et al., 2021). Many results obtained from clinical studies indicated the suitability of digestion, absorption, metabolism and distribution of postbiotics (Asif et al., 2023; Gao et al., 2019; Shenderov 2013; Singh et al., 2018), and these characteristics are suitable for the food and pharmaceutical industries (Aghebati-Maleki et al., 2021).
Many studies were carried out to reduce the amount of aflatoxins by postbiotics in food model systems (Karazhiyan et al., 2016; Nassar et al., 2018) and culture media (Taheur et al., 2019; Vosough et al., 2014), but, so far, no study is discovered on the systematic review of articles for (1) determining the amount of aflatoxin reduction by postbiotics and (2) identifying and introducing the most effective postbiotic to reduce aflatoxins.
Materials and Methods
Search Strategy
This systematic review was based on the guidelines of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA), with the reports cited in this review being consistent (Moher et al., 2015). In this study, the population, intervention, comparison and outcome (PICO) approach was used to identify terms.
Our study started on 01 September 2022 and continued up to 27 October 2022. Four electronic databases, including Web of Science, Google Scholar, PubMed and Scopus, were used from the beginning till 25 September 2022. The following keywords were used to search databases: “Probiotics” AND “Postbiotics” OR “Non-viable probiotics” OR “Parapsychobiotics” OR “Paraprobiotics” OR “Heat-killed probiotics” OR “Ghost probiotics” OR “Metabiotics” OR “Cell-free supernatant” OR “Biogenic” OR “Tyndallized probiotics” OR “Probiotic lysates” AND “Detoxification” AND “Aflatoxin.” These words along with synonyms or Medical Subject Heading (MeSH) terms were added to the search terms.
Duplicate cases were excluded from the final selection of the study. As this review focused on published papers, gray literature found on Google Scholar was not included. After selecting relevant titles and abstracts, all texts of the selected articles were evaluated. A PRISMA flowchart was used to graphically represent the selection of final articles (Figure 3).
Figure 3. PRISMA flow chart of the systematic review search process.
Inclusion and exclusion criteria
Articles were considered eligible if the full-text articles met the below-mentioned conditions:
-
Full text of articles must be accessible
-
Article must be written in English
-
Articles were peer-reviewed
-
Articles written about the detoxification of aflatoxin by postbiotics
-
Articles about the inhibitory effect of probiotic metabolites or killed probiotics against aflatoxin
-
In vitro articles
-
Articles that considered probiotic bacteria
Articles that did not use postbiotics as an inhibitory agent and dealt with other methods or that document in vivo conditions were excluded from the study. Furthermore, conference abstracts, letters to editors, and non-peer-reviewed studies were also excluded. In this study, two reviewers were used to include the articles. They screened all titles and abstracts independently to find eligible articles. Sometimes, if needed, the third referee was consulted to clear doubts, increase self-confidence and reduce errors. For example, if it was not specified in articles whether postbiotics or probiotics were used, a third reviewer was consulted for clarity.
Data extraction
Two reviewers independently extracted the data using a pre-designed table. The extracted data included article title, year of publication, probiotics type, type of aflatoxin, results, and other relevant findings. The extracted data were analyzed.
Quality assessment
Risk of bias assessment for each article included evaluation in the six areas of research logic: description of methodology, rationale of research, description of results, characterization and testing, overall conclusions, and description of discussion (Khizer et al., 2021).
The current risk of bias framework was adapted from the Cochrane Handbook for Systematic Reviews of Interventions (Tarsilla, 2010). Owing to the lack of a suitable risk assessment tool in this particular field, namely food safety, the evaluation process of each included study was carried out independently by two members of the review team. The final decision was that once any disagreements were resolved in the process, a categorization and binding agreement would be reached.
At the end of the work, a systematic review checklist was used to ensure accuracy of work process (Page et al., 2021).
Results and Discussion
Content extraction
The present paper considered a new field of research that focused on biological compounds derived from probiotics, with a limited number of available studies. The information for this study was extracted from 29 articles in line with the objectives of the present study. It should be noted that the results of this systematic review have limitations, as in some papers, it was not mentioned whether the strain in question was a probiotic or not.
Various factors, such as proving that LABs are probiotics, type of inactivation of microorganisms and extraction methods, and the exact nature of the consumed postbiotic (cell-free supernatant or cells killed by different physical and chemical treatments), were also considered. Therefore, in the future studies, it would be important to determine the aforementioned factors to obtain healthy and anti-toxic effects of the considered postbiotics.
Creation and isolation of postbiotics
As mentioned, probiotics must be alive to exert their health effects, but their viability has created limitations for use and treatment. In recent years, it has been shown that the viability of a probiotic is not related to its beneficial effects. Hence, the focus on the health benefits of microorganisms has shifted from live probiotics to postbiotics. According to different definitions, postbiotics include both nonviable bacteria and substances derived from bacterial metabolism. These side products provide specific physiological effects to host by providing more biological activity (Cicenia et al., 2016), and because postbiotics do not contain live microorganisms, the risks associated with their consumption are minimized. Furthermore, postbiotics can be used in a controlled and standardized manner (Shigwedha, 2014). The problem of dose standardization, which is a significant issue in probiotic production, does not exist in the case of postbiotics. Production methods have been investigated by researchers to produce postbiotics in a controlled, standardized, and efficient manner to enable their analysis and use in food, pharmaceuticals and nutritional programs. Although there are no clear international regulatory standards for postbiotics, in some countries, postbiotics have been included in their national regulations and their products have entered public domain (Gelmetti et al., 2022; Kiran et al., 2023). The use of postbiotics derived from probiotics is a specific, effective, environment-friendly, cheap and safe strategy. Since postbiotics have unique features, such as specific chemical structure, safe and long shelf life, they can be used in a delivery system, such as medicine or beneficial foods, to achieve wellness goals, prevention and treatment of diseases (Shigwedha, 2014; Tomar et al., 2015).
In the food industry, postbiotics are produced using natural and laboratory (physical and chemical) methods. One of the most important methods to produce postbiotics in a natural form is the fermentation process. In this, prebiotic compounds are rendered to microorganisms involved in the fermentation process to produce postbiotics (Izuddin et al., 2018). By using different laboratory methods, postbiotics are produced in a pure form with higher performance (Dunand et al., 2019). To inactivate and destroy bacterial cells, physical methods, such as mechanical breaking, heat, gamma, or ultraviolet radiation, high hydrostatic pressure and ultrasound, or chemical methods, such as inactivation by acid, are used (Fan et al., 2021). These methods change cellular structures and their physiological functions. Bacteria are destroyed by applying these methods and are no longer able to grow, but they still maintain their positive effects (de Almada et al., 2016; Rad et al., 2020b). In order to destroy and inactivate bacterial cells, extraction postbiotics by solvent and sonicator are also used (Amaretti et al., 2013). After this, steps such as centrifugation, dialysis and freeze-drying are also used to increase the efficiency of production and storage of these compounds (Amaretti et al., 2013). Despite the existence of various techniques to obtain postbiotics, it is essential to optimize culture environments and bacterial growth conditions along with using new methods of extraction and identification. After achieving optimal production conditions on a laboratory scale, optimization in industry is recommended to maximize production. Finally, appropriate dose and frequency of use are determined.
Table 1. Postbiotics derived from probiotics to eliminate the toxic effect of AFB1, B2, G1, G2 and M1 in food model systems and culture media.
| Probiotic strains | Treatment for obtaining postbiotics/type postbiotics | Medium kind | Probiotics concentration (CFU/mL) | Exposure time | AFB1removal (%) | References |
|---|---|---|---|---|---|---|
| L. rhamnosus (GG) | Heat-killed | Liquid media | - | 4 h | 81 | El-Nezami et al., 1998a |
| L. rhamnosus (LC-705) | Heat-killed | Liquid media | - | 4 h | 82 | El-Nezami et al., 1998b |
| L. rhamnosus (GG) | Acid-killed | Aqueous solution | 1×109 | - | 75.52 | Ghofrani et al., 2018 |
| L. rhamnosus (GG) | Cell wall | Aqueous solution | 1×109 | - | 75.28 | Ghofrani et al., 2018 |
| E. faecium (EF031) | Heat-killed | Aqueous solution | 1×1010 | 48 h | 37.5 | Topcu et al., 2010 |
| E. faecium (M74) | Heat-killed | Aqueous solution | 1×1010 | 24 h | 30.5 | Topcu et al., 2010 |
| L. plantarum | Heat-killed | Aqueous solution | 1×1010 | 48 h | 60-70 | Damayanti et al., 2017 |
| L. rhamnosus (GG) | Heat-killed | Cotton seed | 1×109 | 24 h | 47 | Vosough et al., 2013 |
| L. rhamnosus (GG) | Acid-killed | Cotton seed | 1×109 | 24 h | 49 | Vosough et al., 2013 |
| L. rhamnosus (GG) | Trypsin-treated | Aqueous solution | - | - | 88 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | ɑ-Chymotrypsin-treated | Aqueous solution | - | - | 56 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | N-glycoproteinase F-treated | Aqueous solution | - | - | 85 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | SDS-treated | Aqueous solution | - | - | 89 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | Urea-treated | Aqueous solution | - | - | 78 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | CaCl2-treated | Aqueous solution | - | - | 49 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | MgCl2-treated | Aqueous solution | - | - | 54 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | EDTA-treated | Aqueous solution | - | - | 49 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | EGTA-treated | Aqueous solution | - | - | 50 | Lahtinen et al., 2004 |
| L. rhamnosus (GG) | Heat-killed | PBS | 1×1010 | - | 85 | Haskard et al., 2000 |
| L. rhamnosus (GG) | Acid-killed | PBS | 1×1010 | - | 91 | Haskard et al., 2000 |
| L. rhamnosus (GG) | Acid-killed | Yoghurt | 2.1×109 | 24 h | 77.9 | Karazhiyan et al., 2016 |
| L. rhamnosus (GG) | Heat-killed | Yoghurt | 2.1×109 | 24 h | 72.5 | Karazhiyan et al., 2016 |
| L. rhamnosus (GG) | Ultrasound-treated | Yoghurt | 2.1×109 | 24 h | 78.9 | Karazhiyan et al., 2016 |
| L. plantarum (C88) | Heat-killed | Aqueous solution | 1×1010 | - | 50–60 | Huang et al., 2017 |
| B. licheniformis (CFR1) | CFS1 | PBS2 | - | 72 h | 93.75 | Rao et al., 2017 |
| E. faecium (MF4) | Heat-killed | PBS | 1×108 | 24 h | 70–80 | Fernandez et al., 2015 |
| E. faecium(GJ40) | Heat-killed | PBS | 1×109 | 24 h | 80–90 | Fernandez et al., 2015 |
| L. rhamnosus (GG) | Heat-killed | PBS | 109 | 12 h | 49 | Vosough et al., 2014 |
| L. rhamnosus (GG) | Acid-killed | PBS | 109 | 12 h | 50 | Vosough et al., 2014 |
| L. brevis | CFS | PBS | - | 7 d | 80–90 | Taheur et al., 2019 |
| P.pentosaceus | CFS | PBS | - | 7 d | 30–40 | Taheur et al., 2019 |
| L. kefiri | CFS | PBS | - | 7 d | 91.87 | Taheur et al., 2019 |
| B. subtilis (ANSB060) | CFS | Aqueous solution | 108 | 72 h | 78.7 | Gao et al., 2011 |
| L. reuteri (NRRL14171) | EPS | Aqueous solution | 1-3×1010 | 4 h | <5 | Hernandez-Mendoza et al., 2009 |
| L. reuteri (NRRL14171) | Protoplasts | Aqueous solution | 1-3×1010 | 4 h | <10 | Hernandez-Mendoza et al., 2009 |
| L. reuteri (NRRL14171) | Spheroplasts | Aqueous solution | 1-3×1010 | 4 h | 30–40 | Hernandez-Mendoza et al., 2009 |
| L. reuteri (NRRL14171) | Bacterial cell wall | Aqueous solution | 1-3×1010 | 4 h | 60–70 | Hernandez-Mendoza et al., 2009 |
| L. reuteri (NRRL14171) | Teichoic acid-deficient bacteria | Aqueous solution | 1-3×1010 | 4 h | 20 | Hernandez-Mendoza et al., 2009 |
| L. reuteri (NRRL14171) | Bacteria treated with the polycation | Aqueous solution | 1-3×1010 | 4 h | 40–50 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | EPS3 | Aqueous solution | 1-3×1010 | 4 h | <10 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | Protoplasts | Aqueous solution | 1-3×1010 | 4 h | 10–20 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | Spheroplasts | Aqueous solution | 1-3×1010 | 4 h | 50–60 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | Bacterial cell wall | Aqueous solution | 1-3×1010 | 4 h | 60–70 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | Teichoic acid-deficient bacteria | Aqueous solution | 1-3×1010 | 4 h | 20–30 | Hernandez-Mendoza et al., 2009 |
| L. casei (Shirota) | Bacteria treated with the polycation | Aqueous solution | 1-3×1010 | 4 h | 40–50 | Hernandez-Mendoza et al., 2009 |
| L. rhamnosus (RC007) | CFS | GIT4simulation | 1×108 | 48 h | 80.40 | Fochesato et al., 2019 |
| L. paracasei (KC39) | Acid-killed | Milk | - | - | 69.56 | Fahim et al., 2021 |
| L. acidophilus(ATCC 20552) | Heat-killed | PBS | 108 | 1 h | 73.8 | Elsanhoty et al., 2013 |
| L. rhamnosus (TISTR 541) | Heat-killed | PBS | 108 | 1 h | 84.7 | Elsanhoty et al., 2013 |
| L. sanfranciscensis (DSM20451) | Heat-killed | PBS | 108 | 1 h | 44.5 | Elsanhoty et al., 2013 |
| B. angulatum (DSMZ 20098) | Heat-killed | PBS | 108 | 1 h | 69.5 | Elsanhoty et al., 2013 |
| L. rhamnosus (GG) | Heat-killed | PBS | 2×109 | 2 h | 47.76 | Balsini et al., 2021 |
| B.amyloliquefaciens (WF2020) | CFS | Liquid culture | - | 96 h | 37.16 | Chen et al., 2022b |
| L. kefiri | CFS | Liquid media | 108 | 7 d | 97.22 | Taheura et al., 2021 |
| L. kunkeei | Heat-killed | PBS | - | - | 54.56 | Ebrahimi et al., 2021 |
| L. casei (Shirota) | Acid-killed | PBS | 109 | 72 h | 62 | Ondiek et al., 2022 |
| L. brevis | CFS | PBS | - | 7 d | 60–70 | Taheur et al., 2019 |
| P. pentosaceus | CFS | PBS | - | 7 d | 30–40 | Taheur et al., 2019 |
| L. kefiri | CFS | PBS | - | 7 d | 95.27 | Taheur et al., 2019 |
| L. acidophilus(ATCC 20552) | Heat-killed | PBS | 108 | 1 h | 69.9 | Elsanhoty et al., 2013 |
| L. rhamnosus (TISTR 541) | Heat-killed | PBS | 108 | 1 h | 80.1 | Elsanhoty et al., 2013 |
| L. sanfranciscensis (DSM20451) | Heat-killed | PBS | 108 | 1 h | 35.4 | Elsanhoty et al., 2013 |
| B. angulatum (DSMZ 20098) | Heat-killed | PBS | 108 | 1 h | 62.9 | Elsanhoty et al., 2013 |
| L. kefiri | CFS | Liquid media | 108 | 7 d | 95.27 | Taheura et al., 2021 |
| L. kefiri | Heat-killed | PBS | - | - | 39.57 | Ebrahimi et al., 2021 |
| B. subtilis (ANSB060) | CFS | Aqueous solution | 108 | 72 h | 61 | Gao et al., 2011 |
| L. acidophilus (ATCC 20552) | Heat-killed | PBS | 108 | 1 h | 71.7 | Elsanhoty et al., 2013 |
| L. rhamnosus (TISTR 541) | Heat-killed | PBS | 108 | 1 h | 86.2 | Elsanhoty et al., 2013 |
| L. sanfranciscensis (DSM20451) | Heat-killed | PBS | 108 | 1 h | 41.4 | Elsanhoty et al., 2013 |
| B. angulatum (DSMZ 20098) | Heat-killed | PBS | 108 | 1 h | 66.8 | Elsanhoty et al., 2013 |
| L. kefiri | Heat-killed | PBS | - | - | 42.48 | Ebrahimi et al., 2021 |
| L. acidophilus (ATCC 20552) | Heat-killed | PBS | 108 | 1 h | 67.9 | Elsanhoty et al., 2013 |
| L. rhamnosus (TISTR 541) | Heat-killed | PBS | 108 | 1 h | 81.4 | Elsanhoty et al., 2013 |
| L. sanfranciscensis (DSM20451) | Heat-killed | PBS | 108 | 1 h | 33.4 | Elsanhoty et al., 2013 |
| B. angulatum (DSMZ 20098) | Heat-killed | PBS | 108 | 1 h | 61.7 | Elsanhoty et al., 2013 |
| L. kefiri | Heat-killed | PBS | - | - | 20.52 | Ebrahimi et al., 2021 |
| B. subtilis (ANSB060) | CFS | Aqueous solution | 108 | 72 h | 75 | Gao et al., 2011 |
| E. faecium | Heat-killed | Milk | 7 | 28 d | 78.4 | Sarlak et al., 2017 |
| L. acidophilus (ATCC 20552) | Heat-killed | PBS | - | 24 h | 43.9 | Elsanhoty et al., 2014 |
| L. rhamnosus (TISTR 541) | Heat-killed | PBS | - | 24 h | 46.2 | Elsanhoty et al., 2014 |
| L.plantrium | Heat-killed | PBS | - | 24 h | 41.3 | Elsanhoty et al., 2014 |
| B. angulatum (DSMZ 20098) | Heat-killed | PBS | - | 24 h | 46.4 | Elsanhoty et al., 2014 |
| P. acidilactici (RC005) | CFS | Milk | - | 48 h | 22 | Martinez et al., 2019 |
| P. pentosaceus (RC006) | CFS | Milk | - | 48 h | 91 | Martinez et al., 2019 |
| L. rhamnosus (RC007) | CFS | Milk | - | 48 h | 28 | Martinez et al., 2019 |
| K. marxianus (VM003) | CFS | Milk | - | 48 h | 100 | Martinez et al., 2019 |
| L. Plantarum | Heat-killed | PBS | 1×109 | 72 h | 51.7 | Abdelmotilib et al., 2018 |
| L. acidophilus | Heat-killed | PBS | 1×109 | 72 h | 67.92 | Abdelmotilib et al., 2018 |
| B. bifidum | Heat-killed | PBS | 1×109 | 72 h | 70.62 | Abdelmotilib et al., 2018 |
| K. lactis | Heat-killed | PBS | 1×109 | 72 h | 69.14 | Abdelmotilib et al., 2018 |
| CPYS-NV5 | Heat-killed | PBS | 1×109 | 72 h | 87.92 | Abdelmotilib et al., 2018 |
| L. rhamnosus (ATCC 7469) | Heat-killed | UHT milk | 1010 | 24 h | 12.3 | Nassar et al., 2018) |
| L. acidophilus (DSMZ 20079) | Heat-killed | UHT milk | 1010 | 24 h | 27.3 | Nassar et al., 2018 |
| L. rhamnosus (ATCC 7469) | Bacteriocin | UHT milk | 1010 | 24 h | 77.3 | Nassar et al., 2018 |
| L. acidophilus (DSMZ 20079) | Bacteriocin | UHT milk | 1010 | 24 h | 72.6 | Nassar et al., 2018 |
| L. paracasei (108) | Heat-killed | PBS | 108–109 | 24 h | 78.7 | Cruz et al., 2020 |
| L. plantarum (49) | Heat-killed | PBS | 108–109 | 24 h | 76.6 | Cruz et al., 2020 |
| L. fermentum (111) | Heat-killed | PBS | 108–109 | 24 h | 78.3 | Cruz et al., 2020 |
1Cell-free supernatant
2Phosphate-buffered salin
3Exopolysaccharides
4Gastrointestinal tract
5Combination of nonviable strains (B. bifidum + L. acidophilus + L. Plantarum + S. cerevisiae + k. lactis)
It is important to note that postbiotics, unlike probiotics, do not add new species to intestinal microbiota, but like prebiotics, they develop the existing probiotics in the digestive system (Homayouni-Rad et al., 2020, 2021). Therefore, it is necessary to carry out a comprehensive investigation for safety, optimization of different extraction methods, identification of the chemical composition of postbiotics, and the physiological role of postbiotics in the biological system to facilitate and use postbiotic compounds in food formulations.
The mechanism of action of postbiotics on the detoxification of aflatoxins
In the biological strategy, detoxification is done through a microbe or non-pathogenic enzymes and/or their catabolic pathways not only to reduce/remove but also to suppress toxins (Afshar et al., 2020). One of these non-pathogenic microbes is probiotics, considered as safe (GRAS) and having health benefits (Afshar et al., 2020). Probiotics can counteract the toxic effects of aflatoxins by the following three mechanisms: (1) Possibility of probiotics competing with aflatoxigenic mold strains to occupy space or an ecological niche, use available nutrients, and reduce the biosynthesis of aflatoxins (probiotics), (2) degradation of aflatoxins by enzymes (postbiotics), and (3) bind aflatoxins to the cell wall of probiotic strains and prevent its intestinal absorption (postbiotics) (Zhao et al., 2016). In general, postbiotics can be classified into different groups based on their biological structure (lipid, protein, carbohydrate, etc.) (Thorakkattu et al., 2022). Different types of postbiotics and their action in detoxification of aflatoxins are shown in Table 2.
Table 2. Different types of postbiotics and their action in detoxification of aflatoxins.
| Different types of postbiotics | Method of detoxification of aflatoxin | References |
|---|---|---|
| Supernatant without cells | Degradation | Chen et al., 2022a |
| Exopolysaccharide | Binding, adsorption | Taheur et al., 2017 |
| Cell wall parts (cell surface proteins, teichoic acids and peptidoglycan) | Binding, adsorption | Khosravi-Darani et al., 2020; Liew et al., 2018; Zolfaghari et al., 2020 |
| Short-chain fatty acids | Degradation | Śliżewska and Smulikowska, 2011 |
| Vitamins | - | - |
| Cell lysate | Binding, adsorption | Lahtinen et al., 2004 |
Some studies reported that probiotics can produce metabolites, such as organic acids, bacteriocins and even hydrogen peroxide, that inhibit production of aflatoxins and also interfere in the biosynthesis of aflatoxins (Gerbaldo et al., 2012; Gourama and Bullerman 1995).
Probiotics can even remove the produced aflatoxins using biodegradation or biosorption mechanisms (Afshar et al., 2020). For removal of aflatoxins, biodegradation is a permanent process but takes a longer period than biological absorption. Biodegradation is done by extracellular or intracellular enzymes (Zhao et al., 2016). Aflatoxins are decomposed by different mechanisms, such as epoxidation, hydroxylation, dehydrogenation, and reduction, depending on the decomposing agent (Martinez et al., 2019). Double furan ring and coumarin structure are common in the chemical structure of aflatoxins (Adebo et al., 2017; Dalié et al., 2010). During microbial degradation, microbial catabolic pathways and specific enzymes present in cell-free supernatant influence coumarin conformational changes and difuran ring cleavage in aflatoxins (Adebo et al., 2017; Dalié et al., 2010).
Degradation or enzymatic conversion is the biological breakdown of mycotoxins into less or nontoxic compounds; only certain cells of bacteria and fungi are effective in this change (Chlebicz and Śliżewska 2020). In biodegradation, aflatoxins are converted into other undesirable metabolites, such as aflatoxicol (AFL), which is possibly harmful to health (Ji et al., 2016). Therefore, it appears that biological absorption is a more suitable method than biological degradation.
Biological absorption is a short-term process, and the formed complex is unstable and depends on the number of absorption sites and the affinity of aflatoxins to these sites (Adebo et al., 2017; Afshar et al., 2020; Luo et al., 2018). In biological absorption, an unspecified physical connection to the cell wall takes place, and aflatoxins are removed by physical adhesion. Aflatoxins bind to the carbohydrate components of the cell wall. Covalent bindings or metabolic degradation is not used in this mechanism. Therefore, nonviable bacteria also can bind and remove aflatoxin (Bueno et al., 2007; Hernandez-Mendoza et al., 2010).
Probiotic bacteria, similar to other Gram-positive bacteria, have a complex arrangement of macromolecules. The wall with a special structure contains compounds such as peptidoglycan (PG) sacculus that surround cytoplasmic membrane and other glycopolymers, such as teichoic acids (TAs) or polysaccharides (PSs) and proteins that decorate PG. Each of these compounds has a different function, for example, the TA unit is divided into two groups: (1) wall teichoic acids (WTAs), which are covalently linked to the PG molecule and (2) lipoteichoic acids (LTAs), which are anchored in cytoplasmic membrane with a glycolipid moiety (Chapot-Chartier and Kulakauskas 2014). WTA composition may constitute half of the total dry weight of the cell wall in certain bacterial species and has high hydrophobicity ( Chapot-Chartier and Kulakauskas 2014; Swoboda et al., 2010).
Exopolysaccharide) EPS(, teichoic or lipoteichoic acid, and peptidoglycan are the three main forms of carbohydrates present in the cell wall. As reported by Lahtinen (2004), there was no evidence of the involvement of EPS, cell wall proteins, Ca2+, or Mg2+ in AFB1 binding. The study confirmed that EPS is rejected as a possible component of binding AFB1, and cell wall peptidoglycan appeared as the most likely carbohydrate component effective in the binding process (Lahtinen et al., 2004). Teichoic acid also has a little effect on binding of mycotoxin (Zhao et al., 2016).
Aflatoxins bind to bacterial cell wall through weak non-covalent interactions, which occur with hydrophobic pockets on its surfaces, such as peptidoglycan and other cell wall polysaccharides, or their compounds that are strongly associated with bacterial peptidoglycan (Figure 4) (Fouad et al., 2021). These non-covalent interactions include van der Waals interaction, electrostatic interactions, and hydrogen bonds. Van der Waals interaction was weaker, compared to hydrogen bonds, but since their large numbers maintain the stability of molecular structure, they play a major role in AFB1 binding (Fouad et al., 2021). Hydrogen bonds account for a small fraction of binding energy; however, hydrogen bonding may be important in the binding of aflatoxins ( Haskard et al., 2001; Lahtinen et al., 2004).
Figure 4. Schematic mechanism of aflatoxin detoxification in the cell wall of Gram-positive bacteria, such as probiotics.
With the two parameters mentioned below, the efficiency of a microorganism as an aflatoxin binder can be determined using the following formula:
Efficiency = (M × Keq),
where M is the number of binding sites in the cell wall of the microorganism, which is different in each microorganism, and Keq is the equilibrium constant of the reaction (Bueno et al., 2007).
According to the above-mentioned formula, differences in the structure of the cell wall and cell membrane of bacteria lead to differences between strains in the absorption of aflatoxins ( Kosztik et al., 2020; Pop et al., 2022; Serrano-Niño et al., 2013).
However, it seems that more studies are needed to identify the precise structures of the cell wall responsible for binding mechanism. Therefore, after recognizing the involved components, their functions are defined in vivo experiments and can be used as a method to detoxify food and feed (Lahtinen et al., 2004).
Necessary conditions to increase the removal of aflatoxins in postbiotics
In addition to the strain, some experimental factors also affect the capacity of aflatoxin removal by probiotics. Since the viability of probiotics at low pH conditions is reduced by passing through the stomach, the ability of nonviable probiotics to detoxify aflatoxins is an important topic to explore (Hamidi et al., 2013; Topcu et al., 2010). It has been shown that bacterial viability is not a prerequisite for aflatoxin detoxification (Sarlak et al., 2017). Using killed probiotics is a safe method, because according to the literature, nonviable bacteria can remove more aflatoxin than live bacteria without causing any problems to humans (Vosough et al., 2013). Bovo et al. (2013), Kabak and Var (2008) and Pierides et al. (2000) found that by inactivating the lactic acid bacteria by heat, their ability to remove AFM1 significantly increases in PBS and milk, compared to live bacteria. El-Nezami et al. (1998a) reported that heat-treated dairy strains had the same removal capacity as live strains. It appears that by applying heat treatment, polysaccharides and peptidoglycans are strongly affected, resulting in denaturation of proteins, and the hydrophobic nature of the surface increases, or Maillard reaction products are formed. These changes cause aflatoxins to bind to bacterial cell wall and plasmatic membrane components, which are not available if the cell wall remains intact (Haskard et al., 2001).
Acid treatment has also been shown to significantly increase the ability of bacteria to bind to AFB1 (El-Nezami et al., 1998a; Peltonen et al., 2001). Haskard et al. (2001) also stated that in most cases, acid treatment is more effective than heat treatment. Several studies showed that acid treatment of yeast or LAB had the highest aflatoxin absorption, compared to live and heat-treated microorganisms (Haskard et al., 2001; Hegazy et al., 2011; Rahaei et al., 2010). A study conducted in 1998 by El-Nezami and colleagues (1998a) stated that acid pretreatment increases the binding ability of nonviable LABs. Acidic pretreatment may lead to the cleavage of amine bonds in peptides and proteins, yielding peptides and amino acids (El-Nezami, et al., 1998a). Haskard et al. (2001) reported that acid treatment may affect cell wall components, such as peptidoglycan and polysaccharide, releasing monomers and further fragmentation into aldehydes after breaking glycosidic bonds. Haskard et al. (2000) pointed out that acid-treated LAB may show more hydrophobic binding sites for aflatoxins due to protein denaturation, thus expecting hydrophobic interactions. Although the peptidoglycan layer in probiotics is very thick, the acid may reduce thickness and cross-links, leading to larger and more pores being available (Moghaddam et al., 2019). Finally, aflatoxins are easily and quickly bound by the components of cytoplasmic membrane under acidic conditions (Bejaoui et al., 2004).
Pretreatments of probiotic bacteria increases the efficiency of probiotics as adsorbents of aflatoxins because they cause protein denaturation, release some components and increase pore size, and, probably, affect change in charge distribution and hydrophobic nature of bacterial surface (Ahlberg et al., 2015; Karazhiyan et al., 2016). As a result, it allows aflatoxins to bind to the cell wall or its associated components because the available aflatoxin binding sites are increased (El-Nezami et al., 2002). Haskard et al. (2000) expected salts to affect the surface charge of the bacteria, followed by electrostatic interactions, and the effect of salt addition was greater for nonviable cells, as demonstrated by Balsini et al. (2021).
In one study, treatment with sodium dodecyl sulfate caused a significant increase in the ability of the studied strain to bind AFB1. Under sodium dodecyl sulfate, the protein was denatured and the cell wall composed of peptidoglycan was separated (Maragos, 2008).
Treatments such as divalent cations (Ca2+ and Mg2+), chelators (EDTA and EGTA) as well as ultrasound and enzyme treatments, including various proteases, were ineffective to bind aflatoxins (Lahtinen et al., 2004).
Yeasts have been shown to have a mechanism similar to probiotic bacteria in binding AFB1 as a tool for detoxification (Shetty and Jespersen, 2006; Wu et al., 2009). The cell walls of probiotic yeasts (e.g., Saccharomyces cerevisiae) also contain polysaccharides, such as glucan and mannan, which provide different mechanisms to bind with toxins (ionic, hydrogen, and hydrophobic bonds) (Saeidi and Safari, 2010). In probiotic yeasts, heat causes the denaturation of proteins or the formation of Maillard reaction products in the cell wall. By dissolving some of the mannoproteins in the cell wall, the permeability of the wall increases and leads to increased access to the hidden places of the cell wall as observed in probiotic bacteria. In addition, acid conditions act by affecting polysaccharides, converting monomers and breaking them into aldehydes, and some of the reactions may be intracellular in these acidic conditions (Karazhiyan et al., 2016; Oliveira et al., 2013; Rahaie et al., 2010). As a result, aflatoxin are easily and rapidly bound by yeasts under acidic or thermal treatment (Zhong et al., 2022).
Thermal treatments are the main and most common methods for preparing postbiotics, and these include pasteurization, sterilization, Ohmic heating, radio frequency heating and microwave heating. The most common deactivation mechanism of these methods is the thermal effect, but due to different modes of heat generation, they show different deactivation efficiencies. Thermal treatments are low-cost, simple, and common methods for microbial inactivation, but they can cause irreparable damage to some physiologically active substances in microbial cells, such as proteins and polypeptides. Therefore, it is necessary to use more efficient and practical microbial inactivation methods, such as nonthermal treatments (Zhong et al., 2022). In general, heat treatments mainly damage the cell membrane, break the nucleic acid, inactivate essential enzymes, denature protein and polymerizes such as the ribosome. However, the choice of temperature and time for postbiotic preparation depends on the type of microorganism, culture conditions, growth stage, PH and water activity (Zhong et al., 2022). Some microorganisms inactivated by heat treatment showed various beneficial effects for aflatoxin removal (Abdelmotilib et al., 2018; Montaseri et al., 2014).
Removal and release of aflatoxin is a fast and reversible physical process without any chemical change in aflatoxins. The amount of toxin that is removed depends on various factors, such as probiotic/postbiotic strain concentration, treatment time, amount of cell wall components obtained from viable or nonviable bacteria, toxin characteristics, such as toxin concentration and toxin type, and pH of the environment (Ahlberg et al., 2015; Pfliegler et al., 2015). For example, it appears that 108–109 CFU/mL of viable or nonviable probiotic LAB are required to achieve a 50% aflatoxin removal (Bovo et al., 2013; El-Nezami et al., 1998a). Maximum and minimum binding capacity was observed at 1010 CFU/mL and 107 CFU/mL, respectively (Ismail et al., 2017). Maximum binding occurs within the first few minutes of the reaction. The quick connection process is between 0 and 4 h. When the maximum amount of aflatoxins was removed by probiotics strains, no further reduction occurred. Lee et al. (2003) reported that more the aflatoxin is adsorbed by bacterial cells, more the time the adsorbed aflatoxin molecules would remain on the cell surface of bacteria (Lee et al., 2003). In some studies, it was shown that the mixture of probiotics, bacteria or bacteria with yeast had a better effect in reducing aflatoxins (Florina et al., 2018; Mokoena et al., 2006; Okeke et al., 2015). Owing to different food matrices, physicochemical characteristics, and treatment conditions, the rate of aflatoxin reduction is different.
Probiotic strains or postbiotics can bind with aflatoxins and form a probiotic–aflatoxin or postbiotic–aflatoxin complex. The stability of probiotic/postbiotic and aflatoxin complexes depends on strain, treatment and environmental conditions. Since the binding of aflatoxin to probiotics/postbiotics is weak and partially reversible (Peltonen et al., 2001; Pizzolitto et al., 2012; Topcu et al., 2010), in vivo experiments appear as necessary, because aflatoxins may be released from the gastrointestinal tract, resulting in negative health consequences. The possibility of reversibility is 27.8–94.4%, depending on the strain (Moghaddam et al., 2019). It is determined that LAB shows a significant adhesion to intestinal cells, which lose this characteristic by binding to aflatoxins, so the bacteria–-aflatoxin complex is quickly excreted in the digestive system (Gratz et al., 2010).
As mentioned in the detoxification of aflatoxins by probiotics/postbiotics, the process depends on the strain, irrespective of viable or nonviable probiotic (Topcu et al., 2010). The studies conducted on the anti-aflatoxin effect of postbiotics included B1, B2, G1,and G2, the most effective of which are listed in Table 3.
Table 3. The most effective postbiotics.
| Probiotic strains | Type of postbiotics | Type of AF | AF removal (%) | References |
|---|---|---|---|---|
| L. kefiri(MH107106) | CFS | B1 | 97.22 | Taheura et al., 2021 |
| L. kefiri(MH107106) | CFS | B2 | 95.27 | Taheura et al., 2021 |
| L. rhamnosus (TISTR 541) | Heat-killed | G1 | 86.2 | Elsanhoty et al., 2013 |
| L. rhamnosus (TISTR 541) | Heat-killed | G2 | 81.4 | Elsanhoty et al., 2013 |
| P. pentosaceus (RC006) | CFS | M1 | 91 | Martinez et al., 2019 |
Summarizing the previous table, postbiotics derived from probiotics can remove aflatoxins with a mean of 80%. Based on our review, studies have been done more on AFB1, followed by AFM1, probably because of the greater importance of these two aflatoxins. Strain L. rhamnosus (GG) has been mostly used in the literature, and studies have obtained different results, which could be due to food matrix, type of medium, physicochemical characteristics, and different treatment conditions. Among the food items, milk and dairy products are used maximum.
As already reported, after comparing the studies, we concluded that the acid-treated strain was more effective than the heat-treated strain. We also found that cell-free supernatant was very effective in removing aflatoxins. In fact, cell-free supernatant contains active ingredients and bacteriocin (Tenea and Barrigas, 2018), and probably the presence of several compounds makes it effective. It seems that the simultaneous use of several postbiotics derived from different strains can remove aflatoxins in large amounts, resulting in food safety. The result obtained from this study supports the development of future studies for the formulation of biological additives that could be used in functional foods.
Probiotics, prebiotics, synbiotics and postbiotics are novel concepts. However, biological detoxification technology is currently far from complete, and the determination and purification of metabolites is incomplete in many cases. In the future studies, the determination of various factors, such as the proof of LAB being probiotic, types of inactivation of microorganisms and extraction methods, and the exact nature of postbiotic (cell-free supernatant or cells killed by different physical and chemical treatments), to identify health effects and toxicity are essential. More studies are necessary to accurately identify and understand complex relationships between these concepts; this would help to understand the pathway of their mechanism of action. In addition, more research is needed to reveal the dose, time of detoxification, and usage of new preparations to maximize prevention and beneficial effects to reduce toxins on humans. In addition, in vivo, systematic review studies are also needed to support the health claims of postbiotic functional foods. At the same time, a big challenge exists, such as transferring scientific knowledge to business applications, which are a bridge between science and industry. Therefore, it would take a long time to develop biological additives to realize large-scale industrialization (Saeidi and Safari, 2010).
Author contributions
S. Zamanpour and B. Shokrollahi Yancheshmeh independently searched databases and extracted data individually. Any disagreement was resolved by consulting SM Noori, M. Hashemi and A. Afshari. Finally, the full text was written by S. Zamanpour, B. Shokrollahi Yancheshmeh, M. Hashemi and A. Afshari. Text editing was done by SM Noori.
Acknowledgments
We would like to thank the Nutrition Department of Mashhad University of Medical Sciences, Mashhad, Iran, for helping us in this study.
Conflict of Interest
There was no conflict of interest between authors.
REFERENCES
Abbasi, A., Aghebati-Maleki, L. and Homayouni-Rad, A. 2021a. The promising biological role of postbiotics derived from probiotic Lactobacillus species in reproductive health. Crit Rev Food Sci Nutr. 1–13. 10.1080/10408398.2021.1935701
Abbasi, A., Sheykhsaran, E. and Kafil, H.S. 2021b. Postbiotics: Science, Technology and Applications. Bentham Science, Sharjah, UAE. 10.2174/97816810883891210101
Abdelmotilib, N., Hamad, G., Elderea, H., Salem, E. and Sohaimy, S. 2018. Aflatoxin M1 reduction in milk by a novel combination of probiotic bacterial and yeast strains. Eur J Nutr Food Safety. 8(2): 83–99. 10.9734/EJNFS/2018/39486
Abriouel, H., Muñoz, M.d.C.C., Lerma, L.L., Montoro, B.P., Bockelmann, W., Pichner, R., Kabisch, J., Cho, G-S., Franz, C.M.A.P. and Gálvez, A. 2015. New insights in antibiotic resistance of lactobacillus species from fermented foods. Food Res Int. 78: 465–481. 10.1016/j.foodres.2015.09.016
Adebo, O.A., Njobeh, P.B., Gbashi, S., Nwinyi, O.C. and Mavumengwana, V. 2017. Review on microbial degradation of aflatoxins. Crit Rev Food Sci Nutr. 57(15): 3208–3217. 10.1080/10408398.2015.1106440
Afshar, P., Shokrzadeh, M., Raeisi, S.N., Saraei, A.G.-H. and Nasiraii, L.R. 2020. Aflatoxins biodetoxification strategies based on probiotic bacteria. Toxicon 178: 50–58. 10.1016/j.toxicon.2020.02.007
Aghebati-Maleki, L., Hasannezhad, P., Abbasi, A. and Khani, N. 2021. Antibacterial, antiviral, antioxidant, and anticancer activities of postbiotics: a review of mechanisms and therapeutic perspectives. Biointerface Res Appl Chem. 12: 2629–2645. 10.33263/BRIAC122.26292645
Ahlberg, S.H., Joutsjoki, V. and Korhonen, H.J. 2015. Potential of lactic acid bacteria in aflatoxin risk mitigation. Int J Food Microbiol. 207: 87–102. 10.1016/j.ijfoodmicro.2015.04.042
Ahmadi, E. 2020. Potential public health risk due to consumption of contaminated bovine milk with aflatoxin M1 and Coxiella burnetii in the west of Iran. Int J Dairy Technol. 73(3): 479–485. 10.1111/1471-0307.12687
Alshannaq, A. and Yu, J.-H. 2017. Occurrence, toxicity, and analysis of major mycotoxins in food. Int J Environ Res Public Health 14 (6): 632. 10.3390/ijerph14060632
Amaretti, A., Nunzio, M.D., Pompei, A., Raimondi, S., Rossi, M. and Bordoni, A. 2013. Antioxidant properties of potentially probiotic bacteria: in vitro and in vivo activities. Appl Microbiol Biotechnol. 97(2): 809–817. 10.1007/s00253-012-4241-7
Ambadoyiannis, G., Hatzikamari, M., Litopoulou-Tzanetaki, E. and Tzanetakis, N. 2004. Probiotic and technological properties of enterococci isolates from infants and cheese. Food Biotechnol. 18(3): 307–325. 10.1081/FBT-200035024
Armando, M.R., R.P. Pizzolitto., C.A. Dogi., A. Cristofolini., C. Merkis., V. Poloni., A.M. Dalcero and L.R. Cavaglieri. 2012. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J Appl Microbiol. 113(2): 256–264. 10.1111/j.1365-2672.2012.05331.x
Asif, A., Afzaal, M., Shahid, H., Saeed, F., A. Ahmed., Y. Abbas Shah., A. Ejaz., S. Ghani., H. Ateeq and M.R. Khan. 2023. Probing the functional and therapeutic properties of postbiotics in relation to their industrial application. Food Sci. Nutr. 10.1002/fsn3.3465
Balsini, M.S., Dovom, M.R.E., Kadkhodaee, R., Najafi, M.B.H. and Yavarmanesh, M. 2021. Effect of digestion and thermal processing on the stability of microbial cell-aflatoxin B1 complex. LWT 142: 110994. 10.1016/j.lwt.2021.110994
Battacone, G., Nudda, A., Cannas, A., Borlino, A.C., Bomboi, G. and Pulina, G. 2003. Excretion of aflatoxin M1 in milk of dairy ewes treated with different doses of aflatoxin B1. J Dairy Sci. 86(8): 2667–2675. 10.3168/jds.S0022-0302(03)73862-4
Bejaoui, H., Mathieu, F., Taillandier, P. and Lebrihi, A. 2004. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J Appl Microbiol. 97(5): 1038–1044. 10.1111/j.1365-2672.2004.02385.x
Bovo, F., Corassin, C.H., Rosim, R.E. and de Oliveira, C.A.F. 2013. Efficiency of lactic acid bacteria strains for decontamination of aflatoxin M1 in phosphate buffer saline solution and in skimmed milk. Food Bioprocess Technol. 6(8): 2230–2234. 10.1007/s11947-011-0770-9
Bueno, D.J., Casale, C.H., Pizzolitto, T.P., Salvano, M.A. and Oliver, G. 2007. Physical adsorption of aflatoxin B1 by lactic acid bacteria and Saccharomyces cerevisiae: a theoretical model. J Food Protect. 70(9): 2148–2154. 10.4315/0362-028X-70.9.2148
Chartier, C., Pierre, M. and Kulakauskas, S. 2006. Aflatoxin M1 absorption and cytotoxicity on human intestinal in vitro model. Toxicon. 47(4): 409–415. 10.1016/j.toxicon.2005.12.003
Chapot-Chartier, Marie-Pierre and Kulakauskas, S. 2014. Cell wall structure and function in lactic acid bacteria. Microbial Cell Factories 13(1): 1–23. 10.1186/1475-2859-13-S1-S9
Chen, G., Fang, Q., Liao, Z., Xu, C., Liang, Z., Liu, T., Zhong, Q., et al. 2022a. Detoxification of aflatoxin B1 by a potential probiotic bacillus amyloliquefaciens WF2020. Front Microbiol. 13: 891091. 10.3389/fmicb.2022.891091
Chen, G., Fang, Q., Liao, Z., Xu, C., Liang, Z., Liu, T., Zhong, Q., et al. 2022b. Detoxification of aflatoxin B1 by a potential probiotic bacillus amyloliquefaciens WF2020. Front Microbiol. 13. 10.3389/fmicb.2022.891091
Chlebicz, A. and Śliżewska, K. 2020. In vitro detoxification of aflatoxin B1, deoxynivalenol, fumonisins, T-2 toxin and zearalenone by probiotic bacteria from genus Lactobacillus and Saccharomyces cerevisiae yeast. Probiot Antimicrob Prot. 12(1): 289–301. 10.1007/s12602-018-9512-x
Cicenia, A., Santangelo, F., Gambardella, L., Pallotta, L., Iebba, V., Scirocco, A. et al. 2016. Protective role of postbiotic mediators secreted by Lactobacillus rhamnosus GG versus lipopolysaccharide-induced damage in human colonic smooth muscle cells. J Clin Gastroenterol. 50: S140–S144. 10.1097/MCG.0000000000000681
Cruz, P.O., de Matos, C.J., Nascimento, Y.M., Tavares, J.F., de Souza, E.L. and Magalhães, HIF.. 2020. Efficacy of potentially probiotic fruit-derived Lactobacillus fermentum, L. paracasei and L. plantarum to remove aflatoxin M1 in vitro. Toxins. 13(1): 4. 10.3390/toxins13010004
Dalié, D.K.D., Deschamps, A.M. and Richard-Forget, F. 2010. Lactic acid bacteria–potential for control of mould growth and mycotoxins: a review. Food Control. 21(4): 370–380. 10.1016/j.foodcont.2009.07.011
Damayanti, E., Istiqomah, L., Saragih, JE. and Purwoko, T. 2017. Characterization of lactic acid bacteria as poultry probiotic candidates with aflatoxin B1 binding activities. In: IOP Conference Series: Earth and Environmental Science. 10.1088/1755-1315/101/1/012030
Darwish, W.S., Ikenaka, Y., Shouta, M.M.N. and Ishizuka, M. 2014. An overview on mycotoxin contamination of foods in Africa. J Vet Med Sci. 76(6): 789–797. 10.1292/jvms.13-0563
de Almada, C.N., Almada, C.N., Martinez, R.C.R. and Sant’Ana, A.S. 2016. Paraprobiotics: evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci Technol. 58: 96–114. 10.1016/j.tifs.2016.09.011
Dhanasekaran, D., Shanmugapriya, S., Thajuddin, N. and Panneerselvam, A. 2011. Aflatoxins and aflatoxicosis in human and animals. Aflat Biochem Mol Biol. 10(22717): 221–254. 10.5772/22717
Dunand, E., Burns, P., Binetti, A., Bergamini, C., Peralta, G.H., Forzani, L., Reinheimer, J. and Vinderola, G. 2019. Postbiotics produced at laboratory and industrial level as potential functional food ingredients with the capacity to protect mice against Salmonella infection. J Appl Microbiol. 127(1): 219–229. 10.1111/jam.14276
Ebrahimi, M., Sadeghi, A., Rahimi, D., Purabdolah, H. and Shahryari, S. 2021. Postbiotic and anti-aflatoxigenic capabilities of Lactobacillus kunkeei as the potential probiotic LAB isolated from the natural honey. Probiot Antimicrob Prot. 13(2): 343–355. 10.1007/s12602-020-09697-w
El-Nezami, H., Kankaanpaa, P., Salminen, S. and Ahokas, J. 1998a. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem Toxicol. 36(4): 321–326. 10.1016/S0278-6915(97)00160-9
El-Nezami, H., Kankaanpaa, P., Salminen, S. and Ahokas, J. 1998b. Physicochemical alterations enhance the ability of dairy strains of lactic acid bacteria to remove aflatoxin from contaminated media. J Food Protect. 61(4): 466–468. 10.4315/0362-028X-61.4.466
El-Nezami, H., Polychronaki, N., Salminen, S. and Mykkaünen, H. 2002. Binding rather than metabolism may explain the interaction of two food-grade Lactobacillus strains with zearalenone and its derivative ɑ́-zearalenol. Appl Environ Microbiol. 68(7): 3545–3549. 10.1128/AEM.68.7.3545-3549.2002
Elsanhoty, R.M., Ramadan, M.W., El-Gohery, S.S., Abol-Ela, M.F. and Azeke, M.A. 2013. Ability of selected microorganisms for removing aflatoxins in vitro and fate of aflatoxins in contaminated wheat during baladi bread baking. Food Control 33(1): 287–292. 10.1016/j.foodcont.2013.03.002
Elsanhoty, R.M., Salam, S.A., Ramadan, M.F. and Badr, F.H. 2014. Detoxification of aflatoxin M1 in yoghurt using probiotics and lactic acid bacteria. Food Control 43: 129–134. 10.1016/j.foodcont.2014.03.002
Fahim, K.M., Badr, A.N., Shehata, M.G., Hassanen, E.I. and Ahmed, L.I. 2021. Innovative application of postbiotics, parabiotics and encapsulated Lactobacillus plantarum RM1 and Lactobacillus paracasei KC39 for detoxification of aflatoxin M1 in milk powder. J Dairy Res. 88(4): 429–435. 10.1017/S002202992100090X
Fan, K., Wu, J. and Chen, L. 2021. Ultrasound and its combined application in the improvement of microbial and physicochemical quality of fruits and vegetables: a review. Ultrasonics Sonochem. 80: 105838. 10.1016/j.ultsonch.2021.105838
Fernandez J., Guillermina, M., Dalcero, A.M. and Magnoli, C.E. 2015. In vitro aflatoxin B1 binding capacity by two Enterococcus faecium strains isolated from healthy dog faeces. J Appl Microbiol. 118(3): 574–582. 10.1111/jam.12726
Florina, R., Sofia, P., Lia, R., Antoanela, C. and Butnariu, M. 2018. Effect of some probiotic bacteria on the reduction of aflatoxin B1 production in stored arabica coffee beans. Int. Multidiscipl Sci GeoConf SGEM. 18(6.4): 135–142. 10.5593/sgem2018V/6.4/S08.018
Fochesato, A., Silvia, D.C., Poloni, V., Galvagno, M.A., Dogi, C.A. and Cavaglieri, L.R. 2019. Aflatoxin B1 adsorption/desorption dynamics in the presence of Lactobacillus rhamnosus RC 007 in a gastrointestinal tract-simulated model. J Appl Microbiol. 126(1): 223–229. 10.1111/jam.14101
Food and Agriculture Organization (FAO) 2003. Alimentarius Codex 2003. FAO, Rome, Italy.
Fouad, M.T., El-Shenawy, M. and El-Desouky, T.A. 2021. Efficiency of selected lactic acid bacteria isolated from some dairy products on aflatoxin [B. sub. 1] and Ochratoxin A. J Pure Appl Microbiol. 15(1): 312–320. 10.22207/JPAM.15.1.24
Gao, J., Li, Y., Wan, Y., Hu, Y., Liu, L., Yang, S. et al. 2019. A novel post-biotic from Lactobacillus rhamnosus GG with a-beneficial effect on intestinal barrier function. Front Microbiol. 10: 477. 10.3389/fmicb.2019.00477
Gao, X., Ma, Q., Zhao, L., Lei, Y., Shan, Y. and Ji, C. 2011. Isolation of Bacillus subtilis: screening for aflatoxins B1, M1 and G1 detoxification. Eur Food Res Technol. 232(6): 957–962. 10.1007/s00217-011-1463-3
Gelmetti, C., Rigoni, C., Cantù, A.M., Agolzer, A., Agrusa, A., Brena, M. et al. 2022. Topical prebiotics/-postbiotics and PRURISCORE validation in atopic dermatitis. International study of 396 patients. J Dermatol Treat. 1–8. 10.1080/09546634.2022.2131703
Gerbaldo, G.A., Barberis, C., Pascual, L., Dalcero, A. and Barberis, L. 2012. Antifungal activity of two Lactobacillus strains with potential probiotic properties. FEMS Microbiol Let. 332(1): 27–33. 10.1111/j.1574-6968.2012.02570.x
Ghofrani, T., Damoon, H.K., Golian, A. and Heravi, R.H. 2018. In vitro binding potentials of bentonite, yeast cell wall and lactic acid bacteria for aflatoxin B1 and ochratoxin A. Iran J Toxicol. 12(2): 7–13. 10.29252/arakmu.12.2.7
Gourama, H. and Bullerman, L.B. 1995. Inhibition of growth and aflatoxin production of Aspergillus flavus by Lactobacillus species. J Food Protect. 58(11): 1249–1256. 10.4315/0362-028X-58.11.1249
Gratz, S.W., Mykkanen, H. and El-Nezami, H.S. 2010. Probiotics and gut health: a special focus on liver diseases. World J Gastro-enterol (WJG). 16(4): 403. 10.3748/wjg.v16.i4.403
Hamidi, A., Mirnejad, R., Yahaghi, E., Behnod, V., Mirhosseini, A., Amani, S., Sattari, S. and Darian, E.K. 2013. The aflatoxin B1 isolating potential of two lactic acid bacteria. Asian Pacific J Trop Biomed. 3(9): 732–736. 10.1016/S2221-1691(13)60147-1
Haskard, C.A., El-Nezami, H.S., Kankaanpää, P.E., Salminen, S. and Ahokas, J.T. 2001. Surface binding of aflatoxin B1 by lactic acid bacteria. Applied Environ Microbiol. 67(7): 3086–3091. 10.1128/AEM.67.7.3086-3091.2001
Haskard, C., Binnion, C. and Ahokas, J. 2000. Factors affecting the sequestration of aflatoxin by Lactobacillus rhamnosus strain GG. Chem Biol Interactions. 128(1): 39–49. 10.1016/S0009-2797(00)00186-1
Hegazy, E.M., Sadek, Z.I., El-Shafei, K. and El-Khalek, A.B.A. 2011. Aflatoxins binding by Saccharomyces cerevisiae and S. boulardii in functional cereal based ice-cream. Life Sci J (Acta Zhengzhou University Overseas Ed). 8(4): 75–81.
Hernandez-Mendoza, A., Garcia, H.S. and Steele, J.L. 2009a. Screening of Lactobacillus casei strains for their ability to bind aflatoxin B1. Food Chem Toxicol. 47(6): 1064–1068. 10.1016/j.fct.2009.01.042
Hernandez-Mendoza, A., Guzman-De-Peña, D., González-Córdova, A.F., Vallejo-Córdoba, B. and Garcia, H.G. 2010. In vivo assessment of the potential protective effect of Lactobacillus casei Shirota against aflatoxin B1. Dairy Sci Technol. 90(6): 729–740. 10.1051/dst/2010030
Hernandez-Mendoza, A., Guzman-de-Peña, D. and Garcia, H.S. 2009b. Key role of teichoic acids on aflatoxin B1 binding by probiotic bacteria. J Appl Microbiol. 107(2): 395–403. 10.1111/j.1365-2672.2009.04217.x
Homayouni-Rad, A., Fathi-Zavoshti, H., Douroud, N., Shahbazi, N. and Abbas, A. 2020. Evaluating the role of postbiotics as a new generation of probiotics in health and diseases. J Ardabil Univ Med Sci. 19(4): 381–399. 10.29252/jarums.19.4.381
Homayouni-Rad, A., Oroojzadeh, P. and Abbasi, A. 2021. The effect of yeast kluyveromyces marxianus as a probiotic on the microbiological and sensorial properties of set yoghurt during refrigerated storage. J Ardabil Univ Med Sci. 20(2): 254–268. 10.52547/jarums.20.2.254
Huang, Li., Duan, C., Zhao, Y., Gao, L., Niu, C., Xu, J. and Li, S. 2017. Reduction of aflatoxin B1 toxicity by Lactobacillus plantarum C88: a potential probiotic strain isolated from Chinese traditional fermented food tofu. PloS One. 12(1): e0170109. 10.1371/journal.pone.0170109
Ismail, A., Levin, R.E., Riaz, M., Akhtar, S., Gong, Y.Y. and de Oliveira, C.A.F. 2017. Effect of different microbial concentrations on binding of aflatoxin M1 and stability testing. Food Control. 73: 492–496. 10.1016/j.foodcont.2016.08.040
Izuddin, W.I., Loh, T.C., Samsudin, A.A. and Foo, H.L. 2018. In vitro study of postbiotics from Lactobacillus plantarum RG14 on rumen fermentation and microbial population. Revista Brasileira de Zootecnia 47. 10.1590/rbz4720170255
Ji, C., Fan, F. and Zhao, L. 2016. Review on biological degradation of mycotoxins. Animal Nutr. 2(3): 127–133. 10.1016/j.aninu.2016.07.003
Kabak, B. and Var, I. 2008. Factors affecting the removal of aflatoxin M1 from food model by Lactobacillus and Bifidobacterium strains. J Environ Sci Health B. 43(7): 617–624. 10.1080/03601230802234740
Karazhiyan, H., Mehraban, S.M., Karazhyan, R., Mehrzad, A. and Haghighi, E. 2016. Ability of different treatments of Saccharomyces cerevisiae to surface bind aflatoxin M1 in yoghurt.
Khizer, Z., Sadia, A., Sharma, R., Farhaj, S., Nirwan, S.J., Kakadia, P.G., Hussain, T., Yousaf, A.M., Shahzad, Y. and Conway, B.R. 2021. Drug delivery approaches for managing overactive bladder (Oab): a systematic review. Pharmaceuticals. 14(5): 409. 10.3390/ph14050409
Khosravi-Darani, K., Zoghi, A., Jazayeri, S. and da Cruz, A.G. 2020. Decontamination of aflatoxins with a focus on aflatoxin B1 by probiotic bacteria and yeasts: a review. J Microbiol Biotechnol Food Sci. 10(3): 424–435. 10.15414/jmbfs.2020.10.3.424-435
Kim, M.J., Ku, S., Kim, S.Y., Lee, H.H., Jin, H., Kang, S., Li, R. et al. 2018. Safety evaluations of Bifidobacterium bifidum BGN4 and Bifidobacterium longum BORI. Int J Mol Sci. 19(5): 1422. 10.3390/ijms19051422
Kiran, F., Demirhan, H.K., Haliscelik, O. and Zatari, D. 2023. Metabolic profiles of Weissella spp. postbiotics with-anti-microbial and anti-oxidant effects. J Inf Develop Count. 17(04): 507–517. 10.3855/jidc.16921
Kosztik, J., Mörtl, M., Székács, A., Kukolya, J. and Bata-Vidács, I. 2020. Aflatoxin B1 and sterigmatocystin binding potential of lactobacilli. Toxins 12(12): 756. 10.3390/toxins12120756
Kumar, P., Mahato, D.K., Kamle, M., Mohanta, T.K. and Kang, S.G. 2017. Aflatoxins: a global concern for food safety, human health and their management. Front Microbiol. 7: 2170. 10.3389/fmicb.2016.02170
Laciakova, A.N.N.A., Cicooova, P., Mate, D. and Laciak, V. 2008. Aflatoxins and possibilities for their biological detoxification. Medycyna Weterynaryjna 64(3): 276–279.
Lahtinen, S.J., Haskard, C.A., Ouwehand, A.C., Salminen, S.J. and Ahokas, J.T. 2004. Binding of aflatoxin B1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Additives Contam. 21(2): 158–164. 10.1080/02652030310001639521
Lee, Y.K., El-Nezami, H., Haskard, C.A., Gratz, S., Puong, K.Y., Salminen, S. and Mykkänen, H. 2003. Kinetics of adsorption and desorption of aflatoxin B1 by viable and nonviable bacteria. J Food Protect. 66(3): 426–430. 10.4315/0362-028X-66.3.426
Liew, W.-P.-P., Nurul-Adilah, Z., Than, L.T.L. and Mohd-Redzwan, S. 2018. The binding efficiency and interaction of Lactobacillus casei Shirota toward aflatoxin B1. Front Microbiol. 9: 1503. 10.3389/fmicb.2018.01503
Liong, M.-T. 2008. Safety of probiotics: translocation and-infection. Nutr Rev. 66(4): 192–202. 10.1111/j.1753-4887.2008.00024.x
Lizárraga-Paulín, E.G., Moreno-Martínez, E. and Miranda-Castro, S.P. 2011. Aflatoxins and their impact on human and animal health: an emerging problem. Aflatoxins Biochem Mol Biol. 13: 255–262. 10.5772/26196
Luo, Y., Liu, X. and Li, J. 2018. Updating techniques on controlling mycotoxins–a review. Food Control 89: 123–132. 10.1016/j.foodcont.2018.01.016
Fashandi, H.M., Abbasi, R. and Khaneghah, A.M. 2018. The detoxification of aflatoxin M1 by Lactobacillus acidophilus and Bifidobacterium spp.: a review. J Food Proces Preserv. 42(9): e13704. 10.1111/jfpp.13704
Maragos, C.M. 2008. Extraction of aflatoxins B1 and G1 from maize by using aqueous sodium dodecyl sulfate. J AOAC Int. 91(4): 762–767. 10.1093/jaoac/91.4.762
Marchese, S., Polo A., Ariano, A., Velotto, A., Costantini, S. and Severino, L. 2018. Aflatoxin B1 and M1: biological properties and their involvement in cancer development. Toxins 10(6): 214. 10.3390/toxins10060214
Martinez, M.P., Magnoli, A.P., Pereyra, M.L.G. and Cavaglieri, L. 2019. Probiotic bacteria and yeasts adsorb aflatoxin M1 in milk and degrade it to less toxic AFM1-metabolites. Toxicon 172: 1–7. 10.1016/j.toxicon.2019.09.020
Martins, M.L., Martins, H.M. and Bernardo, F. 2001. Aflatoxins in spices marketed in Portugal. Food Addit Contam. 18(4): 315–319. 10.1080/02652030120041
Moghaddam, A.F., Sarlak, Z. and Hosseini, H. 2019. Application of probiotics in aflatoxins risk reduction in foods: a review. Int J Environ Sci Natural Res. 22(3): 88–96. 10.19080/IJESNR.2019.22.556087
Moher, D., Shamseer, L., Clarke, M., Ghersi, D., Liberati, A., Petticrew, M., Shekelle, P. and Stewart, L.A. 2015. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst Rev. 4(1): 1–9. 10.1186/2046-4053-4-1
Mokoena, M.P., Chelule, P.K. and Gqaleni, N. 2006. The toxicity and decreased concentration of aflatoxin B1 in natural lactic acid fermented maize meal. J Appl Microbiol. 100(4): 773–777. 10.1111/j.1365-2672.2006.02881.x
Montaseri, H., Arjmandtalab, S., Dehghanzadeh, G., Karami, S., Razmjoo, M.M., Sayadi, M. and Oryan, A. 2014. Effect of production and storage of probiotic yogurt on aflatoxin M1 residue. J Food Quality Hazards Control 1(1): 7–14.
Moradi, M., Molaei, R. and Guimarães, J.T. 2021. A review on preparation and chemical analysis of postbiotics from lactic acid bacteria. Enzyme Microbial Technol. 143: 109722. 10.1016/j.enzmictec.2020.109722
Nassar, W.S., Elbarbary, H.A., Ibrahim, E., Mohammed, H.A. and Ibrahim, M.I.M. 2018. A new trial in Egypt to detoxify AFM1 in UHT milk by lactobacilli and their bacteriocins. Alexandria J Vet Sci. 59(1): 60–67. 10.5455/ajvs.299013
Oatley, J.T., Rarick, M.D., Ji, G.E. and Linz, J.E. 2000. Binding of aflatoxin B1 to bifidobacteria in vitro. J Food Protect. 63(8): 1133–1136. 10.4315/0362-028X-63.8.1133
Okeke, C.A., Ezekiel, C.N., Nwangburuka, C.C., Sulyok, C., Ezeamagu, C.O., Adeleke, R.A., Dike, S.K. and Krska, R. 2015. Bacterial diversity and mycotoxin reduction during maize fermentation (steeping) for ogi production. Front Microbiol. 6: 1402. 10.3389/fmicb.2015.01402
Oliveira, C.A.F., Bovo, F., Corassin, C.H., Jager, A.V. and Reddy, K.R. 2013. Recent trends in microbiological decontamination of aflatoxins in foodstuffs. Aflatoxins Recent Adv Future Prospects 1: 59–62. 10.5772/51120
Ondiek, W., Wang, Y., Sun, L., Zhou, L., On, S.L.W., Zheng, H. and Ravi, G. 2022. Removal of aflatoxin B1 and T-2 toxin by bacteria isolated from commercially available probiotic dairy foods. Food Sci Technol Int. 28(1): 15–25. 10.1177/1082013220987916
Page, M.J., McKenzie, J.E., Bossuyt, P.M., Boutron, I., Hoffmann, T.C., Mulrow, C.D., Shamseer, L. et al. 2021. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. Syst Rev. 10(1): 1–11. 10.1016/j.ijsu.2021.105906
Peltonen, K., El-Nezami, H., Haskard, C., Ahokas, J. and Salminen, S. 2001. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J Dairy Sci. 84(10): 2152–2156. 10.3168/jds.S0022-0302(01)74660-7
Peng, Z., Chen, L., Zhu, Y., Huang, Y., Hu, X., Wu, Q., Nüssler, A.K., Liu, L. and Yang, W. 2018. Current major degradation methods for aflatoxins: A review. Trends Food Sci Technol. 80: 155–166. 10.1016/j.tifs.2018.08.009
Pfliegler, W.P., Pusztahelyi, T. and Pócsi, I. 2015. Mycotoxins–-prevention and decontamination by yeasts. J Basic Microbio. 55(7): 805–818. 10.1002/jobm.201400833
Pickova, D., Ostry, V. and Malir, F. 2021. A recent overview of producers and important dietary sources of aflatoxins. Toxins 13(3): 186. 10.3390/toxins13030186
Pierides, M., El-Nezami, H., Peltonen, K., Salminen, S. and Ahokas, J. 2000. Ability of dairy strains of lactic acid bacteria to bind aflatoxin M1 in a food model. J Food Prot. 63(5): 645–650. 10.4315/0362-028X-63.5.645
Piqué, N., Berlanga, M. and Miñana-Galbis, D. 2019. Health benefits of heat-killed (tyndallized) probiotics: an overview. Int J Mol Sci. 20(10): 2534. 10.3390/ijms20102534
Pizzolitto, R.P., Salvano, M.A. and Dalcero, A.M. 2012. Analysis of fumonisin B1 removal by microorganisms in co-occurrence with aflatoxin B1 and the nature of the binding process. Int J Food Microbiol. 156(3): 214–221. 10.1016/j.ijfoodmicro.2012.03.024
Pop, O.L., Suharoschi, R. and Gabbianelli, R. 2022. Biodetoxification and protective properties of probiotics. Microorganisms 10(7): 1278. 10.3390/microorganisms10071278
Rad, A.H., Abbasi, A., Javadi, A., Pourjafar, H., Javadi, M. and Khaleghi, M. 2020a. Comparing the microbial quality of traditional and industrial yoghurts. Biointerface Res Appl Chem. 10(4): 6020–6025. 10.33263/BRIAC104.020025
Rad, A.H., Abbasi, A., Kafil, H.S. and Ganbarov, K. 2020b. Potential pharmaceutical and food applications of postbiotics: a review. Curr Pharm Biotechnol. 21(15): 1576–1587. 10.2174/1389201021666200516154833
Rad, A.H., Aghebati-Maleki, L., Kafil, H.S. and Abbasi, A. 2021a. Molecular mechanisms of postbiotics in colorectal cancer prevention and treatment. Crit Rev Food Sci Nutr. 61(11): 1787–1803. 10.1080/10408398.2020.1765310
Rad, A.H., Maleki, A.L., Kafil, H.S., Zavoshti, H.F. and Abbasi, A. 2020c. Postbiotics as novel health-promoting ingredients in functional foods. Health Promotion Perspect. 10(1): 3–4. 10.15171/hpp.2020.02
Rad, A.H., Maleki, A.L., Kafil, H.S., Zavoshti, H.F. and Abbasi, A. 2021b. Postbiotics as promising tools for cancer adjuvant therapy. Adv Pharm Bulletin. 11(1): 1. 10.34172/apb.2021.007
Rahaei, S., Razavi, S.H. and Emam, J.Z. 2010. The ability of Saccharomyces cerevisiae strain in aflatoxin reduction in pistachio nuts. 10.1590/S1517-83822010000100014
Rahaie, S., Emam-Djomeh, Z., Razavi, S.H. and Mazaheri, M. 2010. Immobilized Saccharomyces cerevisiae as a potential aflatoxin decontaminating agent in pistachio nuts. Brazil J Microbiol. 41: 82–90. 10.1590/S1517-83822010000100014
Rao, K.R., Vipin, A,V., Hariprasad, P., Appaiah, K.A.A. and Venkateswaran, G.J.F.C. 2017. Biological detoxification of-aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 71: 234–241. 10.1016/j.foodcont.2016.06.040
Saeidi, A.S.L. and Safari, R. 2010. Inhibitory effect of saccharomyces cerevisiae on G1 and G2 aflatoxins in culture media and kilka fish meal.
Salminen, S., Collado, M.C., Endo, A., Hill, C., Lebeer, S., Quigley, E.M.M., Sanders, M.E., Shamir, R., Swann, J.R. and Szajewska, H. 2021. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nature Rev Gastroenterol Hepatol. 18(9): 649–667. 10.1038/s41575-021-00440-6
Sarlak, Z., Rouhi, M., Mohammadi, R., Khaksar, R., Mortazavian, A.M., Sohrabvandi, S. and Garavand, F. 2017. Probiotic biological strategies to decontaminate aflatoxin M1 in a traditional Iranian fermented milk drink (Doogh). Food Control 71: 152–159. 10.1016/j.foodcont.2016.06.037
Serrano-Niño, J.C., Cavazos-Garduño, A., Hernandez-Mendoza, A., Applegate, B., Ferruzzi, M.G., Martin-González, M.F.S and García, H.S. 2013. Assessment of probiotic strains ability to reduce the bioaccessibility of aflatoxin M1 in artificially contaminated milk using an in vitro digestive model. Food Control 31(1): 202–207. 10.1016/j.foodcont.2012.09.023
Shenderov, B.A. 2013. Metabiotics: novel idea or natural development of probiotic conception. Microbial Ecol Health Dis. 24(1): 20399. 10.3402/mehd.v24i0.20399
Shetty, P.H. and Jespersen, L. 2006. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci Technol. 17(2): 48–55. 10.1016/j.tifs.2005.10.004
Shigwedha, N. 2014. Probiotical cell fragments (PCFs) as novel nutraceutical ingredients. J Biosci Med. 2(03): 43. 10.4236/jbm.2014.23007
Singh, A., Vishwakarma, V. and Singhal, B. 2018. Metabiotics: the functional metabolic signatures of probiotics: current state-of-art and future research priorities—metabiotics: probiotics effector molecules. Adv Biosc Biotechnol. 9(4): 147–189. 10.4236/abb.2018.94012
Sklan, D., lipper, E., Friedman, A., Shelly, M. and Makovsky, B. 2001. The effect of chronic feeding of diacetoxyscirpenol, T–2 toxin and aflatoxin on performance, health and antibody production in chicks. J Appl Poultry Res. 10(1): 79–85. 10.1093/japr/10.1.79
Śliżewska, K and Smulikowska, S. 2011. Detoxification of aflatoxin B. J Animal Feed Sci. 20: 300–309. 10.22358/jafs/66187/2011
Swoboda, J.G., Campbell, J., Meredith, T.C. and Walker, S. 2010. Wall teichoic acid function, biosynthesis and inhibition. Chembiochem 11(1): 35–45. 10.1002/cbic.200900557
Taheur, F.B., Fedhila, K., Chaieb, K., Kouidhi, B., Bakhrouf, A. and Abrunhosa, L. 2017. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int J Food Microbiol. 251: 1–7. 10.1016/j.ijfoodmicro.2017.03.021
Taheur, F.B., Mansour, C., Kouidhi, B. and Chaieb, K. 2019. Use of lactic acid bacteria for the inhibition of Aspergillus flavus and Aspergillus carbonarius growth and mycotoxin production. Toxicon 166: 15–23. 10.1016/j.toxicon.2019.05.004
Taheura, F.B., Mansoura, C. and Mzoughia, R. 2021. Biocontrol potential of Lactobacillus kefiri probiotic strain against Aspergillus flavus and aflatoxin production in almonds.
Tarsilla, M. 2010. Cochrane handbook for systematic reviews of interventions. J Multidiscipl Eval. 6(14): 142–148. 10.56645/jmde.v6i14.284
Tenea, G.N. and Barrigas, A. 2018. The efficacy of bacteriocin-containing cell-free supernatant from Lactobacillus plantarum Cys5-4 to control pathogenic bacteria growth in artisanal beverages. Int Food Res J. 25(5): 2031–2037.
Thorakkattu, P., Khanashyam, A.C., Shah, K., Babu, K.S., Mundanat, A.S., Deliephan, A., Deokar, G.S., Santivarangkna, C. and Nirmal, N.P. 2022. Postbiotics: current trends in food and pharmaceutical industry. Foods 11(19): 3094. 10.3390/foods11193094
Tomar, S.K., Anand, S., Sharma, P., Sangwan, V. and Mandal, S. 2015. Role of probiotics, prebiotics, synbiotics and postbiotics in inhibition of pathogens. The battle against microbial pathogens: Basic Science, Technological Advances and Educational Programs; Méndez-Vilas, A., Ed: 717–732.
Topcu, A., Bulat, T., Wishah, R. and Boyacı, I.H. 2010. Detoxification of aflatoxin B1 and patulin by Enterococcus faecium strains. Int. J Food Microbiol. 139(3): 202–205. 10.1016/j.ijfoodmicro.2010.03.006
Vinderola, G., Ellen Sanders, M. and Salminen, S. 2022. The concept of postbiotics. Foods 11(8): 1077. 10.3390/foods11081077
Vosough, P.R., Sani, A.M., Sangatash, M.M. and Karazhyan, R. 2013. Assessing the ability of Lactobacillus rhamnosus GG to bind aflatoxin B.
Vosough, P.R., Sani, A.M., Sangatash, M.M. and Karazhyan, R. 2014. In vitro effect of Lactobacillus rhamnosus GG on reduction of aflatoxin B1. Nutr Food Sci.
Walhe, R., Khan, H. and Kumari, S. 2022. From probiotics to postbiotics: key to microbiome and health. In: Microbiome-Gut-Brain Axis. Springer, New York, NY, pp. 367–381. 10.1007/978-981-16-1626-6_18
Wu, Q., Jezkova, A., Yuan, Z., Pavlikova, L., Dohnal, V. and Kuca, K. 2009. Biological degradation of aflatoxins. Drug Metabolism Rev. 41(1): 1–7. 10.1080/03602530802563850
Yu, J., Payne, G.A., Campbell, B.C., Guo, B., Cleveland, T.E., Robens, J.F., Keller, N.P., Bennett, J.W. and Nierman, W.C. 2008. Mycotoxin production and prevention of aflatoxin contamination in food and feed. Aspergilli Genomics Med Aspects Biotechnol Res Methods 457–472. 10.1201/9781420008517.ch27
Zain, M.E. 2011. Impact of mycotoxins on humans and animals. J Saudi Chem Soc. 15(2): 129–144. 10.1016/j.jscs.2010.06.006
Zhao, H., Wang, X., Zhang, J., Zhang, J. and Zhang, B. 2016. The mechanism of Lactobacillus strains for their ability to remove fumonisins B1 and B2. Food Chem Toxicol. 97: 40–46. 10.1016/j.fct.2016.08.028
Zhong, Y., Wang, T., Luo, R., Liu, J., Jin, R. and Peng, X. 2022. Recent advances and potentiality of postbiotics in the food industry: Composition, inactivation methods, current applications in metabolic syndrome and future trends. Crit Rev Food Sci Nutr. 1–25. 10.1080/10408398.2022.2158174
Zhou, G., Chen, Y., Kong, Q., Ma, Y. and Liu, Y. 2017. Detoxification of aflatoxin B1 by Zygosaccharomyces rouxii with solid state fermentation in peanut meal. Toxins 9(1): 42. 10.3390/toxins9010042
Zolfaghari, H., Khezerlou, A., Ehsani, A. and Khosroushahi, A.Y. 2020. Detoxification of aflatoxin B1 by probiotic yeasts and bacteria isolated from dairy products of Iran. Adv Pharm Bulletin 10(3): 482. 10.34172/apb.2020.060
Zychowski, K.E., Hoffmann, A.R., Ly, H.J., Pohlenz, C., Buentello, A., Romoser, A., Gatlin, D.M. and Phillips, T.D. 2013. The effect of aflatoxin-B1 on red drum (Sciaenops ocellatus) and assessment of dietary supplementation of NovaSil for the prevention of aflatoxicosis. Toxins 5(9): 1555–1573. 10.3390/toxins5091555