
Download
PAPER
Health benefits of co-supplementing mealworm protein hydrolysate and cranberry fruit extract
Jae Hong Park1#, Sang In Lee2#, Woo Sung Kwon2, Sungbo Cho3*, In Ho Kim1*
1Department of Animal Resource and Science, Dankook University, Cheonan, Republic of Korea;
2Department of Animal Biotechnology, Kyungpook National University, Sangju, Republic of Korea;
3School of Mongolian Medicine, Inner Mongolia University for Nationalities, Tongliao, People’s Republic of China
Abstract
The demand for valuable protein sources is increasing. The mealworm has been highlighted as a good source of protein. Nevertheless, beneficial effects of mealworm such as the antioxidative and/or anti-inflammatory effects are rarely studied. It is well-known that cranberry fruit has a strong antioxidant effect. The biologically active compounds in mealworm and cranberry could boost the antioxidative and/or anti-inflammatory effects. The current study investigated the interactive effects of mealworm protein hydrolysate (MWPH) and cranberry fruit extract (CFE) in mammals. We evaluated growth performance, relative organ weight, immune responses, antioxidant enzyme activities, blood properties, and fecal microflora. A 2 × 2 factorial experimental design was used. The co-supplementation of MWPH and CFE improved serum glutathione peroxidase. MWPH affected a lower serum IL-1β and fecal Clostridium density. The co-supplementation appeared more effective in terms of good health and potentially the prevention of disease.
Key words: cranberry, extracts, immunity, inflammation, mealworm, microbiota
*Corresponding Authors: Sungbo Cho, School of Mongolian Medicine, Inner Mongolia University for Nationalities, Tongliao 028000, Inner Mongolia Autonomous Region, People’s Republic of China. Email: [email protected]; In Ho Kim, Department of Animal Resource and Science, Dankook University, Cheonan, 31116, Republic of Korea. Email: [email protected]
Received: 25 July 2022; Accepted: 15 November 2022; Published: 2 January 2023
#These authors contributed equally to this work.
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Demand for protein is expected to increase in the future, owing to the increasing global population. Recently, insects have been recognized as the best alternatives to meet protein requirements (Van Huis, 2013). The edible larvae of the common pest insect Tenebrio molitor (yellow mealworm; YMW) distributed worldwide are a good source of protein, fat, vitamins, and minerals (Kim et al., 2014). YMWs contain high-quality protein (Shockley and Dossey, 2014), and contain more essential amino acids than soybeans (Yi et al., 2013). In addition, they have higher unsaturated fatty acid content than meat, and are relatively rich in vitamin A and iron (Rumpold and Schlüter, 2013). Recent studies have reported that mealworms can partially replace soybean and fish meal as a livestock protein feed source, and can improve the growth and productivity of various animals, including chicken and fish (Bovera et al., 2015; Ido et al., 2019). Hence, YMWs are extensively used in livestock feed due to the richness of amino acids and proteins (Hong et al., 2020). The YMW is also attracting attention as a useful raw material for the production of physiological active peptides. Physiologically active peptides are natural antioxidants, generally defined as a peptide with physiological activity and low molecular weight, which helps in their easy absorption into the body (Arihara et al., 2001). Their bioactive peptide contents and chitin were known to possess antioxidant and antimicrobial properties (Di Mattia et al., 2019; Matheswaran et al., 2019). Nevertheless, the antioxidant and antimicrobial effects of YMWs were reported in mostly in vitro studies. A few fish feed research demonstrated the antioxidant enzyme activities in rainbow trout (Henry et al., 2018) and olive flounder (Jeong et al., 2021). There is a limited study regarding the antioxidant effects of YMW dietary supplementation in mammals. Recently, Ringseis et al. (2021) reported that the 10% YMW supplementation to growing pigs did not show a significant alternation in the antioxidant enzymes catalase (CAT), glutathione peroxidase (GPX), and superoxide dismutase (SOD) in the liver and of GPX and SOD in gastrocnemius muscle. Therefore, it is tempting to boost the antioxidant effect of YMW by co-supplementing with natural antioxidants.
Cranberries are a group of evergreen dwarf shrubs in the family Ericaceae, belonging to the subgenus Oxycoccus of the genus Vaccinium. Fresh cranberries, which are red in color and have a sour and sweet taste, are mainly cultivated in the cool climates of northern United States, Canada, and Chile. These berries have several biological functions that help prevent chronic disease in humans (Rupasinghe et al., 2015). Cranberries are also rich in phenolic compounds that exert strong antioxidant effects (Cho et al., 2012). Cranberry fruit extracts (CFE) contain a good source of polyphenols such as proanthocyanidins, flavanols (Vvedenskaya and Vorsa, 2004), and quercetin (Duthie et al., 2006; Zheng and Wang, 2003), which is attributed to the strong antioxidant properties. In addition, cranberries are rich in anthocyanins, water-soluble red pigments of the flavonoid family that exhibit anticancer, antioxidant, and anti-aging effects (Song et al., 2018). So far, the antioxidant and anti-inflammatory effect of CFE in mammals are also limited. Therefore, we hypothesized that the combination of YMW and cranberry as a dietary supplement would improve health in mammals. In the present study, we aimed to verify the in vivo efficacy of a diet supplemented with YMW and CFEs either alone or in combination. Toward this, the effects of feeding mealworm protein hydrolysate (MWPH) and cranberry fruit extract (CFE) on mouse growth, organ weight, COX-2 and NF-kB expression, blood parameters, and fecal microflora were investigated.
Materials and Methods
Ethical endorsement
The experimental protocol and procedures used in this study were approved by the Institutional Animal Care and Use Committee of Dankook University (DK-1-2136).
Preparations of MWPH and CFE
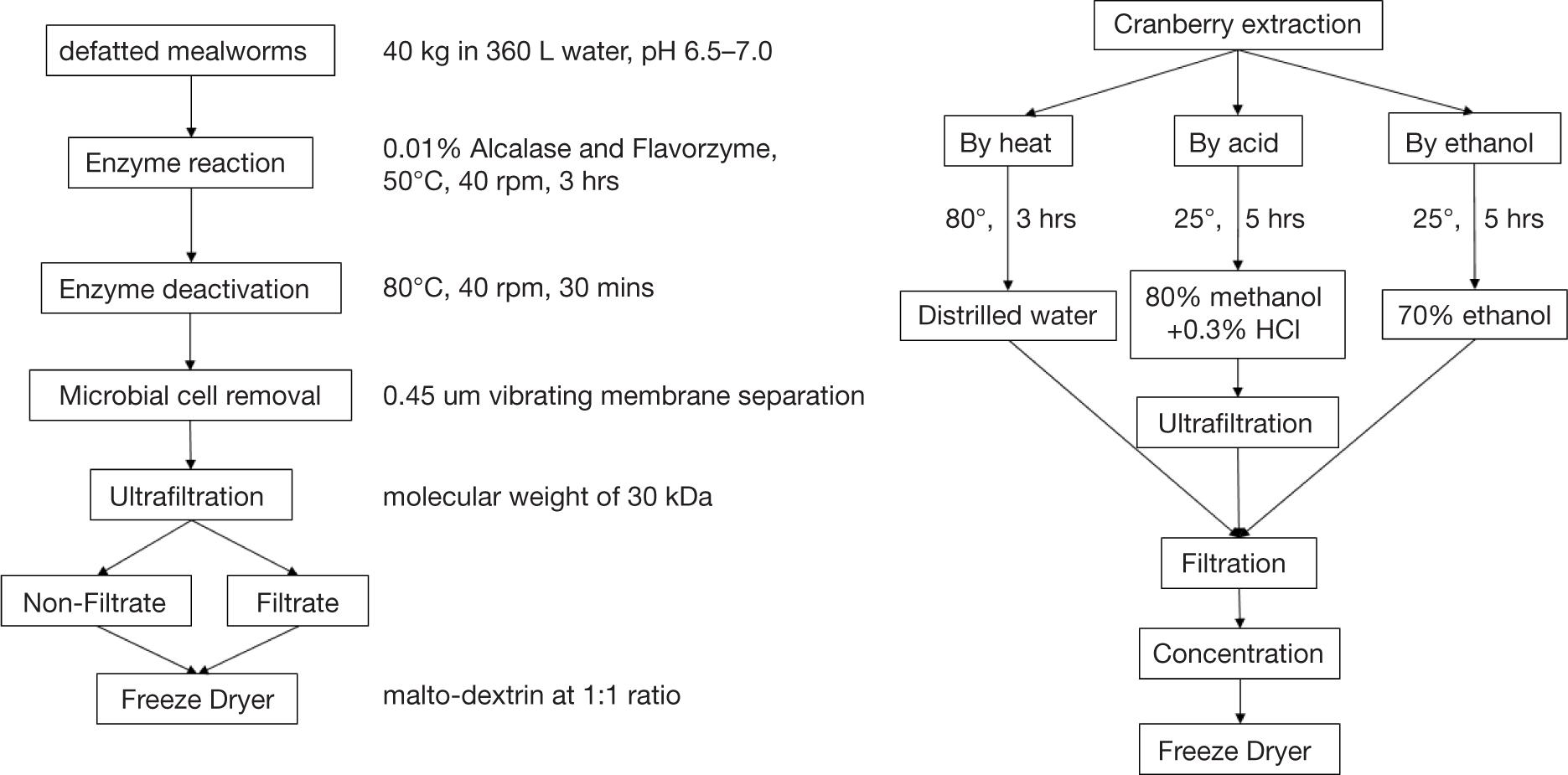
The MWPH and CFE used in this study were purchased from Jeonbuk Institute for Food-Bioindustry (Jeonju, Korea). The MWPH was prepared as follows: 40 kg defatted mealworms were mixed with 360 L water, and the pH was adjusted to 6.5–7.0 using 1 N NaOH. Next, 0.01% (w/w) formulations of each of Alcalase and Flavorzyme were added, and the mixture was hydrolyzed at 50°C for 3 h while stirring at 40 rpm. Subsequently, the mixture was heated at 85°C for 30 min to deactivate the enzymes, followed by centrifugation at 152 rpm under 130 L/h flow rate conditions. Next, microbial cells were removed by following vibrating membrane separation process, after which ultrafiltration was performed to obtain hydrolysate with molecular weight of 100 kDa and 30 kDa. The final hydrolysate was mixed with malto-dextrin at 1:1 ratio and freeze-dried at −60°C before use (Figure 1). The composition of amino acids in unhydrolyzed mealworm and MWPH were analyzed.
Figure 1. The process of cranberry extraction mealworm protein hydrolysate used in the current research.
The CFE was prepared in three different extract methods, namely, hot water, methanol and HCl, and Ethanol. The water extraction was carried at 80°C water for 3 h and, secondly, cranberry fruit was extracted by 80% methanol and 0.3% HCl for 5 h. The third method is as follows: 5-fold volume of 70% ethanol was added to 100 g cranberry fruit and extracted for 5 h using a reflux condenser. This extract was concentrated under reduced pressure at 40 ± 1°C and freeze-dried before use (Figure 1).
Animals and experimental design
A total of 40 mice (BALB/c female inbred, 42 days old) with an initial average body weight of 29.4 ± 0.1 g were purchased from Central Lab. Animal Inc. (Seoul, Korea) and fostered in ventilated cages with a 12 h daylight (06:00–18:00) and 12 h dark cycle. The mice were allowed free access to water and food throughout the study.
Mice were randomly assigned to one of the four experimental treatments (one replication; 10 mice per treatment). The experimental treatments were arranged in a 2 × 2 factorial design with two levels of MWPH (0 and 2%) and two levels of CFE (0 and 400 ppm). CFE was added to the basal diet, while MWPH substituted 2% of the casein protein in the basal diet. Basal diets were prepared based on the American Institution of Nutrition (AIN-93G) guidelines for the nutritional requirements of mice (Table 1).
Table 1. Composition of the basal diets (on as-fed basis).
| Ingredients, g/kg | YMW− | YMW+ | ||
|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | |
| Casein | 200 | 200 | 180 | 180 |
| Sucrose | 100 | 100 | 100 | 100 |
| Dextrose | 132 | 132 | 132 | 132 |
| Corn starch | 398 | 398 | 398 | 398 |
| Cellulose | 50 | 50 | 50 | 50 |
| Soybean oil | 70 | 70 | 70 | 70 |
| Mineral mix | 35 | 35 | 35 | 35 |
| Vitamin mix | 10 | 10 | 10 | 10 |
| L-cystine | 3 | 3 | 3 | 3 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 | 2.5 |
| Yellow mealworm | – | – | 20 | 20 |
| Cranberry fruit extract | – | 0.4 | – | 0.4 |
| Total | 1000.0 | 1000.4 | 1000.0 | 1000.4 |
Sampling and analysis
The amino acid compositions of unhydrolyzed mealworm and MWPH were quantified in an amino acid analyzer (Sykam S-433 D, Sykam GmbH, Eresing, Germany). The amino acid compositions of the samples were compared with those of standards (Sigma Aldrich, St. Louis, MO, USA). The chemical compounds of CFEs from different extract methods were analyzed by HPLC using an Agilent 1100 HPLC system equipped with quaternary pumps, an auto-sampler, and a diode array detector set at 520 nm.
The body weight (BW) of each mouse was measured at the beginning and end of the study. Body weight gain (BWG), feed intake (FI), and feed efficiency (FE) were assessed at the end of the study. On day 42, all mice were placed in metabolic cages and fresh fecal samples were collected. On the same day, blood samples were drawn by using EDTA-treated tubes, stored at 37°C for 2 h, and centrifuged at 3500 rpm for 15 min at 4°C. Then, the supernatants were aspirated and stored at 4°C until further analysis. Serum level of pro-inflammatory cytokines (interleukin-6 [IL-6], IL-1β, and tumor necrosis factor-α [TNF-α]) were measured using a commercial ELISA kit (ELISA MAK; BioLegend, San Diego, CA) and absorbance was measured at 450 nm using a spectrophotometer. Antioxidant enzyme (superoxide dismutase [SOD] and glutamic peroxidase [GPx]) activity was analyzed using specific enzyme assay kits (Cayman Chemical, Michigan, USA). Following blood sampling, mice were anesthetized in chambers saturated with isoflurane and euthanized by cardiac puncture. Kidneys, spleens, and livers were harvested carefully for further analysis, and organ weight was calculated as a percentage of live weight.
DNA extraction
The QIAamp DNA Stool kit (Qiagen) was used to extract DNA from frozen stool samples according to the manufacturer’s instructions. Briefly, the procedure involved lysis of the bacterial cells within the fecal material in ASL buffer, adsorption of impurities that inhibit EX reagent, and purification of the DNA on a spin column. ASL buffer was specially developed to remove inhibitory substances from stool samples. The DNA was eluted in a final volume of 200 µL and stored at −20°C.
The total volume of each amplification reaction mixture was 25 µL, consisting of 1 × TaqMan Universal PCR Master Mix (Applied Biosystems), both primers (each at 300 nM concentration), 200 nM TaqMan MGB probe, 60 ng purified target DNA, and bovine serum albumin at a final concentration of 0.1 µg/µL (New England Biolabs). Amplification (2 min at 50°C, 10 min at 95°C, followed by 45 cycles of 15 s at 95°C and 1 min at 60°C) and detection were carried out on an ABI Prism 7900 sequence detection system (Applied Biosystems). Total bacteria were used as an endogenous control to normalize target gene expression (Table 2).
Table 2. Group-specific primers based on 16S rDNA sequences used in this study.
| Target bacteria | Sequences (5' – 3') | |
|---|---|---|
| Total bacteria | F | ACTCCTACGGGAGGCAG |
| R | GTATTACCGCGGCTGCTG | |
| Bifidobacterium | F | CGCGTCYGGTGTGAAAG |
| R | CCCCACATCCAGCATCCA | |
| Lactobacillus | F | GAGGCAGCAGTAGGGAATCTTC |
| R | GGCCAGTTACTACCTCTATCCTTCTTC | |
| Clostridium | F | GGGGGTTTCAACACCTCC |
| R | GCAAGGGATGTCAAGTGT | |
| Firmicutes | F | GGAGYATGTGGTTTAATTCGAAGCA |
| R | AGCTGACGACAACCATGCAC | |
| Bacteroidetes | F | GAAGGTCCCCCACATTG |
| R | CGCKACTTGGCTGGTTCAG |
Quantitative real-time (qRT)-PCR
To measure the expression of COX-2 and NF-kB, mice were euthanized at the end of the experiment. Kidney samples were frozen in liquid nitrogen and stored at −80°C. Total RNA was isolated from the kidneys using TRIzol (Invitrogen, Carlsbad, CA, USA). For qRT-PCR, total RNA (1 µg) was used as a template for cDNA synthesis performed using the Maxima First-Strand cDNA Synthesis Kit (Life Technologies, Carlsbad, CA, USA). Sequence-specific primers (Table 3) were designed using Primer3 (http://frodo.wi.mit.edu/), and the reaction was performed on the CFX real-time PCR system (Bio-Rad, Hercules, CA, USA) under the following conditions: 94°C for 3 min, followed by 40 cycles of 94°C for 30 s, 59–61°C for 30 s, and 72°C for 30 s. Target gene expression levels were normalized to that of the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase and were calculated using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Table 3. List and sequences of primers used in this study.
| Gene symbol | Description | Accession no. | Forward primer (5' -> 3') | Reverse primer (5' -> 3') |
|---|---|---|---|---|
| Cox-2 | cyclooxygenase-2 | NM011198 | CTCACGAAGG AACTCAGCAC | GGATTGGAAC AGCAAGGATT |
| Nf-Kb1 | nuclear factor of kappa light polypeptide gene enhancer in B cells 1 | NM008689 | GTCTGCCTCTC TCGTCTTCC | CAGTGGGCTG TCTCCAGTAA |
| Gapdh | glyceraldehyde-3-phosphate dehydrogenase | NM001289726 | CGAGACCCCA CTAACATCAA | GGTTCACACC CATCACAAAC |
Statistical analysis
All statistical analyses were performed in a 2 × 2 factorial design using ANOVA tests (SAS Inst. Inc., Cary, NC, US). Statistical significance was set at P < 0.05.
Results
Active compounds and amino acids
The quantity of amino acids in unhydrolyzed mealworm and MWPH were estimated. Both did not contain threonine, histidine, isoleucine, tryptophan, and taurine. The MWPH had higher quantity of amino acids compared to unhydrolyzed mealworm (Table 4).
Table 4. The quantity of amino acid from unhydrolyzed mealworm and mealworm protein hydrolysate (MWPH) (mg/g dry weight).
| Amino acids | Unhydrolyzed mealworm | MWPH | |
|---|---|---|---|
| Essential | Threonine | 0.00 | 0.00 |
| Methionine | 7.49 ± 1.26 | 13.6 ± 1.74 | |
| Lysine | 20.5 ± 1.96 | 34.6 ± 2.68 | |
| Histidine | 0.00 | 0.00 | |
| Isoleucine | 0.00 | 0.00 | |
| Leucine | 0.24 ± 0.06 | 0.93 ± 0.07 | |
| Phenylalanine | 0.16 ± 0.05 | 0.54 ± 0.06 | |
| Tryptophan | 0.00 | 0.00 | |
| Valine | 0.17 ± 0.06 | 0.57 ± 0.05 | |
| Nonessential | Arginine | 0.23 ± 0.04 | 0.63 ± 0.06 |
| Taurine | 0.00 | 0.00 | |
The values represent mean ± SEM, n = 5.
The quantity of active compounds from cranberry fruits was compared based on the three different extract methods. The ethanol methods showed the highest quantity of active compounds; thus, CFE from ethanol was used for the animal trial. The CFE from ethanol methods contained 80.6 and 2.917 µg/mL of total anthocyanin and proanthocyanin, respectively (Table 5).
Table 5. Chemical compounds of cranberry fruit extracts from different extract methods.
| Compounds | Unit | Hot water | Acid | Ethanol |
|---|---|---|---|---|
| Cyanidin 3-Gluciside Chloride | mg/g | 0.000 | 0.125 ± 0.08 | 0.000 |
| Cyanidin 3-Galactoside Chloride | mg/g | 0.226 ± 0.08 | 0.692 ± 0.12 | 0.737 ± 0.17 |
| Cyanidin Chloride | mg/g | 0.022 ± 0.001 | 0.030 ± 0.001 | 0.029 ± 0.001 |
| Proanthocyanin | µg/mL | 0.000 | 0.000 | 2.917 ± 0.07 |
| Total anthocyanin | µg/mL | 0.000 | 0.000 | 80.56 ± 3.45 |
| Total polyphenol | µg/mL | 5.658 ± 0.35 | 9.693 ± 0.42 | 13.14 ± 0.49 |
The values represent mean ± SEM (dry weight), n = 6.
Weight gain, feed intake, and feed efficiency
After 6 weeks, neither MWPH- nor CFE-supplemented diets showed a significant difference in BWG, FI, or FE compared to the control. In addition, there was no significant interactive effect of MWPH and CFE supplementation on BWG, FI, and FE (Table 6).
Table 6. Effects of the MWPH and CFE supplementation on growth performance in mice1.
| Items | MWPH − | MWPH + | SEM | MWPH | CFE | P-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | − | + | − | + | MWPH | CFE | MWPH × CFE | ||
| Initial BW, g | 29.3 | 29.5 | 29.5 | 29.3 | 0.45 | 29.4 | 29.4 | 29.4 | 29.4 | 0.888 | 0.986 | 0.847 |
| Finial BW, g | 32.6 | 33.7 | 32.3 | 32.1 | 0.60 | 33.2 | 32.2 | 32.5 | 33.0 | 0.524 | 0.767 | 0.634 |
| BWG, g | 3.30 | 4.20 | 2.80 | 2.80 | 0.29 | 3.80 | 2.80 | 3.10 | 3.50 | 0.062 | 0.467 | 0.464 |
| FI2, g | 113 | 123 | 126 | 132 | 2.63 | 118 | 129 | 120 | 128 | 0.031 | 0.098 | 0.669 |
| FE3 | 0.029 | 0.034 | 0.022 | 0.021 | 0.004 | 0.032 | 0.020 | 0.026 | 0.028 | 0.066 | 0.752 | 0.505 |
MWPH = mealworm protein hydrolysate, CFE = cranberry fruit extract, + or – = supplemented with or without 2% MWPH and 400 ppm CFE, respectively. BWG, body weight gain; FI, feed intake; FE, feed efficiency.
Organ size
There were no differences in the relative weights of the kidney and liver between mice fed diets with MWPH or CFE and those fed unmodified diets. However, the relative weight of the spleen increased with supplementation of MWPH (P < 0.05). No interactive effects were observed of MWPH and CFE supplementation on the relative organ weights (Table 7).
Table 7. Effects of the MWPH and CFE supplementation on relative organ weight in mice1.
| Items | MWPH − | MWPH + | SEM | MWPH | CFE | P-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | − | + | − | + | MWPH | CFE | MWPH × CFE | ||
| Kidney (R), % | 0.52 | 0.54 | 0.59 | 0.55 | 0.009 | 0.53 | 0.57 | 0.55 | 0.55 | 0.127 | 0.812 | 0.115 |
| Kidney (L), % | 0.53 | 0.53 | 0.61 | 0.55 | 0.01 | 0.53 | 0.58 | 0.57 | 0.54 | 0.156 | 0.382 | 0.266 |
| Spleen, % | 0.25 | 0.24 | 0.27 | 0.32 | 0.01 | 0.25 | 0.30 | 0.26 | 0.28 | 0.015 | 0.461 | 0.109 |
| Liver, % | 4.09 | 4.26 | 4.35 | 4.24 | 0.06 | 4.18 | 4.29 | 4.22 | 4.25 | 0.373 | 0.787 | 0.292 |
MWPH = mealworm protein hydrolysate, CFE = cranberry fruit extract, + or – = supplemented with or without 2% MWPH and 400 ppm CFE, respectively.
COX-2 and NF-kB expression
The expression of COX-2 and NF-kB was not influenced by supplementation with MWPH or CFE. Also, there was no significant interactive effect of MWPH plus CFE supplementation on the expression of COX-2 and NF-kB (Table 8).
Table 8. Effects of the MWPH and CFE supplementation on COX2 and NF-kB expression in mice1.
| Items | MWPH − | MWPH + | SEM | MWPH | CFE | P-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | − | + | − | + | MWPH | CFE | MWPH × CFE | ||
| COX-2 | 1.00 | 0.88 | 1.19 | 1.16 | 0.13 | 0.94 | 1.18 | 1.10 | 1.02 | 0.426 | 0.803 | 0.873 |
| NF-κB | 1.00 | 0.60 | 0.65 | 0.64 | 0.11 | 0.80 | 0.65 | 0.83 | 0.62 | 0.553 | 0.422 | 0.451 |
MWPH = mealworm protein hydrolysate, CFE = cranberry fruit extract, + or – = supplemented with or without 2% MWPH and 400 ppm CFE, respectively. COX-2, cyclooxygenase-2; NF-κB, nuclear factor kappa B.
Serum cytokine and antioxidant enzyme levels
Serum IL-1β concentration was lower in mice fed the diet containing MWPH than in those fed diets lacking MWPH (P < 0.05). The GPx concentration in mice fed diets containing MWPH or CFE was higher than that in mice fed diets lacking MWPH or CFE (P < 0.05). The concentrations of IL-6, TNF-α, and SOD were not influenced by MWPH or CFE supplementation. In addition, there was not an interaction between MWPH and CFE supplementation on serum levels of cytokines and antioxidant enzymes (P < 0.05; Table 9).
Table 9. Effects of the MWPH and CFE supplementation on serum profiles of mice1.
| Items | MWPH − | MWPH + | SEM | MWPH | CFE | P-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | − | + | − | + | MWPH | CFE | MWPH × CFE | ||
| IL-6, pg/mL | 130.4 | 75.73 | 50.82 | 46.30 | 30.3 | 103.1 | 48.60 | 90.6 | 61.0 | 0.418 | 0.657 | 0.706 |
| IL-1β, pg/mL | 54.94 | 65.46 | 33.67 | 28.60 | 4.95 | 60.20 | 31.14 | 44.30 | 47.03 | 0.001 | 0.697 | 0.276 |
| TNF-α, pg/mL | 12.86 | 9.05 | 10.46 | 10.02 | 0.76 | 10.95 | 10.23 | 11.65 | 9.53 | 0.643 | 0.184 | 0.286 |
| SOD, U/mL | 4.50 | 7.27 | 8.36 | 7.62 | 0.78 | 5.89 | 7.99 | 6.43 | 7.44 | 0.199 | 0.525 | 0.277 |
| GPx, U/mL | 498.9 | 612.5 | 936.2 | 1382 | 98.1 | 555.7 | 1159 | 717.6 | 997.4 | 0.001 | 0.012 | 0.103 |
MWPH = mealworm protein hydrolysate, CFE = cranberry fruit extract, and + or – = supplemented with or without 2% MWPH and 400 ppm CFE, respectively. IL-6, interleukin-6; IL-1β, Interleukin-1β; TNF-α, tumor necrosis factor-α, SOD, superoxide dismutase; GPx, glutathione peroxidase.
Fecal bacteria
There were no differences in bacterial counts of Bifidobacterium, Lactobacillus, Bacteroidetes, and Firmicutes in mice fed diets containing MWPH or CFE, but Clostridium counts decreased with the supplementation of MWPH (P < 0.05). There was no significant interactive effect of MWPH and CFE supplementation on fecal bacteria (Table 10).
Table 10. Effects of the MWPH and CFE supplementation on fecal microbiota in mice1.
| Items | MWPH − | MWPH + | SEM | MWPH | CFE | P-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CFE− | CFE+ | CFE− | CFE+ | − | + | − | + | MWPH | CFE | MWPH × CFE | ||
| Bifidobacterium spp., log10 | 9.12 | 9.31 | 9.24 | 9.32 | 0.15 | 9.22 | 9.28 | 9.18 | 9.32 | 0.328 | 0.945 | 0.537 |
| Lactobacillus spp., log10 | 8.25 | 8.48 | 7.79 | 8.20 | 0.13 | 8.36 | 8.00 | 8.02 | 8.35 | 0.205 | 0.258 | 0.749 |
| Clostridium spp., log10 | 9.79 | 9.96 | 8.36 | 9.03 | 0.22 | 9.88 | 8.70 | 9.08 | 9.50 | 0.003 | 0.187 | 0.414 |
| Bacteroidetes, % | 48.4 | 49.4 | 49.5 | 49.2 | 0.32 | 48.9 | 49.4 | 49.0 | 49.3 | 0.539 | 0.664 | 0.381 |
| Firmicutes, % | 51.6 | 50.6 | 50.5 | 50.8 | 0.32 | 51.1 | 50.6 | 51.0 | 50.7 | 0.539 | 0.664 | 0.381 |
MWPH = mealworm protein hydrolysate, CFE = cranberry fruit extract, + or – = supplemented with or without 2% MWPH and 400 ppm CFE, respectively.
Discussion
The study was carried out to investigate the healthy benefits of co-supplementing MWPH and CFE in mice. Organ weight can be an important indicator of pathology, and measurements of the liver, kidneys, and spleen are recognized as indicators of toxicity and immunity in vivo. Since there was no difference in the liver and kidney weights of mice in this study, our data suggest that neither MWPH nor CFE had any harmful effects in the mice. However, MWPH supplementation increased spleen weight in this study. It is generally known that immunopotentiators increase the weight of the thymus and spleen (Chen et al., 2012). Since the spleen is largely composed of immune cells, it can be inferred that MWPH supplementation increases spleen immune cell.
The protein complex NF-κB is involved in inflammatory response regulation, immune modulation, apoptosis, cell proliferation, and epithelial differentiation among other processes. It regulates the expression of various genes and forms the central axis of the intracellular signaling system. The cytokines TNF-α, IL-1, IL-6, and the inducible enzyme COX-2 are regulated by NF-κB (Liu et al., 2017). In a previous in vitro study based on lipopolysaccharide-activated macrophages, mealworm larval protein inhibited the expression of iNOS and COX-2 proteins in a concentration-dependent manner (Seo et al., 2019). However, this effect was not observed in the mice in our study.
Cytokines are relatively small molecular weight immune proteins that play an important role in cell signaling (Arango Duque and Descoteaux, 2014). Cytokines involved in inflammatory processes are classified as either inflammatory or anti-inflammatory. Inflammatory cytokines, such as IL-1β, IL-6, and TNF-α, are involved in immune-related disorders, such as rheumatoid arthritis, atopic dermatitis, and asthma (Girodet et al., 2016; Sato et al., 1999; Turner et al., 2014). Anti-inflammatory cytokines such as IL-10 and TGF-β protect against these diseases by performing functions such as the induction of B cells proliferation and tissue repair (Girodet et al., 2016). In this study, IL-1β levels were significantly lower in the MWPH supplementation group. These results suggest that MWPH exerts anti-inflammatory effects via regulating cytokine expression and may help suppress the conversion to chronic inflammation due to enhanced immune function.
Antioxidants are known to slow aging by scavenging cellular oxidants and active radicals in the body (Kim et al., 2003). Several physiologically active peptides act as natural antioxidants and function in hypertension relief, immune regulation, pain relief, and antibacterial action (Xing et al., 2021). They are usually composed of 3–20 amino acids with a low molecular weight that makes it easy to absorb them into the body (Kim et al., 2013). Previous studies have reported that protein hydrolysate from mealworm larvae (Yu et al., 2017) and Protaetia brevitarsis larvae (Lee et al., 2017) increased antioxidant activity. Therefore, in the present study, the increase in GPx activity in mouse blood can be considered as the effect of physiologically active peptides from MWPH.
Cranberries exhibit high antioxidant activity, which is closely related to flavonoid pigments such as anthocyanins, flavonols, and proanthocyanidins (Brown and Shipley, 2011; Feghali et al., 2012). The antioxidant capacity of proanthocyanidins is reportedly reported to be stronger than that of vitamins C and E and catechins (Ariga, 2004). In this study, GPx levels were higher in mice fed on CFE, which is likely due to the abundant polyphenolic compounds in CFE.
Furthermore, CFE can contribute to gut microbiota regulation. The positive effects of polyphenols derived from many plant sources on the gut microbiota have already been investigated (Anhê et al., 2017; Heyman-Lindén et al., 2016; Roopchand et al., 2015). However, in our study, CFE did not affect fecal microbiota in mice fed on CFE-supplemented diet when compared with mice fed on nonsupplemented diet.
In this study, the number of Clostridium bacteria was significantly lower in the mice fed on MWPH supplemented diet, showing that MWPH supplementation inhibits the growth of harmful microorganisms in the intestine. The outer shell of mealworm larvae is mainly composed of Brown mealworm larvae that develop an exoskeleton during metamorphosis into pupae. It has been reported that chitin can be used as a source of dietary fiber that provides a favorable environment for the growth of beneficial bacteria in the gastrointestinal tract (Hamed et al., 2016; Ringø et al., 2012). We therefore expect that chitin contained in MWPH may have influenced our results.
Conclusions
Combination of MWPH and CFE could be advantageous for boosting anti-inflammation via regulating cytokine activation. The combination significantly reduced the expression of IL-1 as compared to individual supplements. Besides, MWPH-supplemented diet improved the immune function and reduced harmful gut bacteria in mice. The effect of CFE supplementation on mice increased levels of antioxidant enzymes in serum. This study provides preliminary data for the further evaluation of dietary supplementation with MWPH and CFE. In future studies, it is necessary to investigate the relative effects of these two additives at various feed levels.
REFERENCES
Anhê, F.F., Nachbar, R.T., Varin, T.V., Vilela, V., Dudonné, S., Pilon, G., Fournier, M., Lecours, M.-A., Desjardins, Y. and Roy, D., 2017. A polyphenol-rich cranberry extract reverses insulin resistance and hepatic steatosis independently of body weight loss. Molecular Metabolism 6(12): 1563–1573. 10.1016/j.molmet.2017.10.003
Arango Duque, G. and Descoteaux, A., 2014. Macrophage cytokines: involvement in immunity and infectious diseases. Frontiers in Immunology 5: 491. 10.3389/fimmu.2014.00491
Ariga, T., 2004. The antioxidative function, preventive action on disease and utilization of proanthocyanidins. Biofactors 21(1–4): 197–201. 10.1002/biof.552210140
Arihara, K., Nakashima, Y., Mukai, T., Ishikawa, S. and Itoh, M., 2001. Peptide inhibitors for angiotensin I-converting enzyme from enzymatic hydrolysates of porcine skeletal muscle proteins. Meat Science 57(3): 319–324. 10.1016/S0309-1740(00)00108-X
Bovera, F., Piccolo, G., Gasco, L., Marono, S., Loponte, R., Vassalotti, G., Mastellone, V., Lombardi, P., Attia, Y. and Nizza, A., 2015. Yellow mealworm larvae (Tenebrio molitor, L.) as a possible alternative to soybean meal in broiler diets. British Poultry Science 56(5): 569–575. 10.1080/00071668.2015.1080815
Brown, P.N. and Shipley, P.R., 2011. Determination of anthocyanins in cranberry fruit and cranberry fruit products by high--performance liquid chromatography with ultraviolet detection: single-laboratory validation. Journal of AOAC International 94(2): 459–466. 10.1093/jaoac/94.2.459
Chen, Y., Tang, J., Wang, X., Sun, F. and Liang, S., 2012. An immunostimulatory polysaccharide (SCP-IIa) from the fruit of Schisandra chinensis (Turcz.) Baill. International Journal of Biological Macromolecules 50(3): 844–848. 10.1016/j.ijbiomac.2011.11.015
Cho, J.-W., An, T.-H., Lee, S.-Y. and Park, K.W., 2012. Determination of total content of phenolic compounds in Chinese matrimony vine’s accessions. Korean Journal of Crop Science 57(4): 409–417. 10.7740/kjcs.2012.57.4.409
Di Mattia, C., Battista, N., Sacchetti, G. and Serafini, M., 2019. Antioxidant activities in vitro of water and liposoluble extracts obtained by different species of edible insects and invertebrates. Frontiers in Nutrition 6:106. 10.3389/fnut.2019.00106
Duthie, S.J., Jenkinson, A.M., Crozier, A., Mullen, W., Pirie, L., Kyle, J., Yap, L.S., Christen, P. and Duthie, G.G., 2006. The effects of cranberry juice consumption on antioxidant status and biomarkers relating to heart disease and cancer in healthy human volunteers. European Journal of Nutrition 45(2), 113–122. 10.1007/s00394-005-0572-9
Feghali, K., Feldman, M., La, V.D., Santos, J. and Grenier, D., 2012. Cranberry proanthocyanidins: natural weapons against periodontal diseases. Journal of Agricultural and Food Chemistry 60(23): 5728–5735. 10.1021/jf203304v
Girodet, P.-O., Nguyen, D., Mancini, J.D., Hundal, M., Zhou, X., Israel, E. and Cernadas, M., 2016. Alternative macrophage activation is increased in asthma. American Journal of Respiratory Cell and Molecular Biology 55(4): 467–475. 10.1165/rcmb.2015-0295OC
Hamed, I., Özogul, F. and Regenstein, J.M., 2016. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): a review. Trends in Food Science & Technology 48: 40–50. 10.1016/j.tifs.2015.11.007
Henry, M., Gai, F., Enes, P., Peréz-Jiménez, A. and Gasco, L., 2018. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish & Shellfish Immunology 83: 308–313. 10.1016/j.fsi.2018.09.040
Heyman-Lindén, L., Kotowska, D., Sand, E., Bjursell, M., Plaza, M., Turner, C., Holm, C., Fåk, F. and Berger, K., 2016. Lingonberries alter the gut microbiota and prevent low-grade inflammation in high-fat diet fed mice. Food & Nutrition Research 60(1): 29993. 10.3402/fnr.v60.29993
Hong, J., Han, T. and Kim, Y.Y., 2020. Mealworm (Tenebrio molitor Larvae) as an alternative protein source for monogastric animal: a review. Animals 10(11): 2068. 10.3390/ani10112068
Ido, A., Hashizume, A., Ohta, T., Takahashi, T., Miura, C. and Miura, T., 2019. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pargus major). Animals 9(3): 100. 10.3390/ani9030100
Jeong, S.-M., Khosravi, S., Yoon, K.-Y., Kim, K.-W., Lee, B.-J., Hur, S.-W. and Lee, S.-M., 2021. Mealworm, Tenebrio molitor, as a feed ingredient for juvenile olive flounder, Paralichthys olivaceus. Aquaculture Reports 20: 100747. 10.1016/j.aqrep.2021.100747
Kim, N.H., Choi, D.W. and Song, K.B., 2013. Preparation of chicken feather protein hydrolysates and isolation of iron-binding peptides. Korean Journal of Food Preservation 20(3): 435–439. 10.11002/kjfp.2013.20.3.435
Kim, S., Kim, J., Oh, H., Kang, S., Koo, H., Kim, H. and Choi, H., 2014. Feed supplementation of yellow mealworms (Tenebrio molitor L.) improves blood characteristics and meat quality in broiler. Trends in Agriculture & Life Sciences 49: 9–18. 10.29335/tals.2014.49.9
Kim, T.-K., Shin, H.-D. and Lee, Y.-H., 2003. Stabilization of polyphenolic antioxidants using inclusion complexation with cyclodextrin and their utilization as the fresh-food preservative. Korean Journal of Food Science and Technology 35(2): 266–271.
Lee, H.-S., Ryu, H.-J., Song, H.-J. and Lee, S.-O., 2017. Enzymatic preparation and antioxidant activities of protein hydrolysates from Protaetia brevitarsis larvae. Journal of the Korean Society of Food Science and Nutrition 46(10): 1164–1170.
Liu, T., Zhang, L., Joo, D. and Sun, S.-C., 2017. NF-κB signaling in inflammation. Signal Transduction and Targeted Therapy 2(1): 17023. 10.1038/sigtrans.2017.23
Livak, K.J. and Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 25(4): 402–408. 10.1006/meth.2001.1262
Matheswaran, P., Raja, L. and Gani, S., 2019. Antioxidant and anti-inflammatory efficacy of functional proteins obtained from seven edible insects. International Journal of Entomology Research 4: 24–31.
Ringø, E., Zhou, Z., Olsen, R. and Song, S., 2012. Use of chitin and krill in aquaculture–the effect on gut microbiota and the immune system: a review. Aquaculture Nutrition 18(2): 117–131. 10.1111/j.1365-2095.2011.00919.x
Ringseis, R., Peter, L., Gessner, D.K., Meyer, S., Most, E. and Eder, K., 2021. Effect of Tenebrio molitor larvae meal on the antioxidant status and stress response pathways in tissues of growing pigs. Archives of Animal Nutrition 75(4): 237–250. 10.1080/1745039X.2021.1950106
Roopchand, D.E., Carmody, R.N., Kuhn, P., Moskal, K., Rojas-Silva, P., Turnbaugh, P.J. and Raskin, I., 2015. Dietary polyphenols promote growth of the gut bacterium Akkermansia muciniphila and attenuate high-fat diet–induced metabolic syndrome. Diabetes 64(8): 2847–2858. 10.2337/db14-1916
Rumpold, B.A. and Schlüter, O.K., 2013. Nutritional composition and safety aspects of edible insects. Molecular Nutrition & Food Research 57(5): 802–823. 10.1002/mnfr.201200735
Rupasinghe, H., Boehm, M., Sekhon-Loodu, S., Parmar, I., Bors, B. and Jamieson, A.R., 2015. Anti-inflammatory activity of haskap cultivars is polyphenols-dependent. Biomolecules 5(2): 1079–1098. 10.3390/biom5021079
Sato, Y., Ohshima, T. and Kondo, T., 1999. Regulatory role of endogenous interleukin-10 in cutaneous inflammatory response of murine wound healing. Biochemical and Biophysical Research Communications 265(1): 194–199. 10.1006/bbrc.1999.1455
Seo, M., Lee, H.J., Lee, J.H., Baek, M., Kim, I.-W., Kim, S.Y., Hwang, J.-S. and Kim, M., 2019. A study of the anti-inflammatory effect of protein derived from Tenebrio molitor larvae. Journal of Life Science 29(8): 854–860.
Shockley, M. and Dossey, A.T., 2014. Insects for human consumption. In: Mass production of beneficial organisms. Elsevier, pp. 617–652.
Song, Y.S., Kim, M.W., Moon, C., Seo, D.J., Han, Y.S., Jo, Y.H., Noh, M.Y., Park, Y.K., Kim, S.A. and Kim, Y.W., 2018. Extraction of chitin and chitosan from larval exuvium and whole body of edible mealworm, Tenebrio molitor. Entomological Research 48(3): 227–233. 10.1111/1748-5967.12304
Turner, M.D., Nedjai, B., Hurst, T. and Pennington, D.J., 2014. Cytokines and chemokines: at the crossroads of cell signalling and inflammatory disease. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1843(11): 2563–2582. 10.1016/j.bbamcr.2014.05.014
Van Huis, A., 2013. Potential of insects as food and feed in assuring food security. Annual Review of Entomology 58: 563–583. 10.1146/annurev-ento-120811-153704
Vvedenskaya, I.O. and Vorsa, N., 2004. Flavonoid composition over fruit development and maturation in American cranberry, Vaccinium macrocarpon Ait. Plant Science 167(5): 1043–1054. 10.1016/j.plantsci.2004.06.001
Xing, L., Li, G., Toldrá, F. and Zhang, W., 2021. The physiological activity of bioactive peptides obtained from meat and meat by-products. Advances in Food and Nutrition Research No. 97. Elsevier, pp. 147–185.
Yi, L., Lakemond, C.M., Sagis, L.M., Eisner-Schadler, V., van Huis, A. and van Boekel, M.A., 2013. Extraction and-characterisation of protein fractions from five insect species. Food Chemistry 141(4): 3341–3348. 10.1016/j.foodchem.2013.05.115
Yu, M.-H., Lee, H.-S., Cho, H.-R. and Lee, S.-O., 2017. Enzymatic preparation and antioxidant activities of protein hydrolysates from Tenebrio molitor larvae (Mealworm). Journal of the Korean Society of Food Science and Nutrition 46(4): 435–441. 10.3746/jkfn.2017.46.4.435
Zheng, W. and Wang, S.Y., 2003. Oxygen radical absorbing capacity of phenolics in blueberries, cranberries, chokeberries, and lingonberries. Journal of Agricultural and Food Chemistry 51(2): 502–509. 10.1021/jf020728u